«Редактирование генома или генное редактирование — метод генной инженерии, позволяющий вставлять, удалять, модифицировать или заменять последовательности ДНК в живых организмах». Это определение, взятое из «Википедии», кажется на первый взгляд немного громоздким, но довольно точно отражает не только суть, но и потенциал, заложенный в этом виде генного лечения. Нужно лишь немного над ним призадуматься...
В самом деле, сегодня мы многое знаем про генную терапию (ей, например, посвящен этот спецпроект!), причем не только про ее завораживающие возможности, но и про серьезные недостатки. Один из них — необходимость доставки в клетку рабочей копии всего поломанного гена целиком, что не всегда бывает оптимально. Дело в том, что обычно достаточно изменить совсем крошечную часть гена, чтобы он снова заработал; то есть вовсе не обязательно включать в лечебную ДНК весь «джентльменский набор» кодирующих и регуляторных последовательностей.
В этом смысле идеальным решением было бы «чинить» дефектный ген в его естественном положении — прямо в хромосоме, просто заменяя несколько «букв» нуклеотидной последовательности, что сделало бы вмешательство гораздо более аккуратным и точным. Одно из преимуществ тут в том, что продукция целевого белка таким «родным» геном вряд ли «угаснет» со временем (как это иногда случается с доставленными извне полноразмерными генами), что очень важно. Ведь вся идея геномного редактирования (как и любой другой генной терапии) — это одноразовое лечение с долгоиграющим эффектом: замедлением или остановкой прогрессирования заболевания на всю жизнь.
Как гласит философский принцип «бритвы Оккама»: не нужно множить сущности сверх необходимого; и для медицины эта пословица более чем актуальна. Обычно чем менее инвазивно, хирургически аккуратно и точно медицинское вмешательство, тем лучше. Генное редактирование позволяет реализовывать именно такую точечную инженерию, расширяя возможности существующих методов лечения и при этом сводя к минимуму вмешательство в естественные молекулярные процессы . Это обусловливает его недюжинный терапевтический потенциал, делая перспективным и привлекательным биомедицинским методом.
Оговоримся: тут речь о теоретическом преимуществе перед классической генно-заместительной терапией, которое в полной мере (как хочется верить!) еще только будет реализовано. Пока же ошибочное редактирование — одна из основных опасностей такого рода терапий, что вызывает недоверие к ним (об этом ниже еще поговорим). Конечно, такие опасения в небезопасности (стоит признать, небезосновательные) характерны и для «обычной» генной терапии, но пока к ней доверия все-таки немного побольше, ведь уже накоплен некоторый опыт ее использования. Подробнее обо всем многообразии генного лечения узнайте из предыдущих статей этого спецпроекта.
Как работает генное редактирование?
Немного истории
В 1985 году англо-американский генетик и биохимик Оливер Смитис продемонстрировал принципиальную возможность внедрять фрагменты ДНК в клетки млекопитающих. В своих экспериментах Смитис с коллегами использовали ген человеческого β-глобина, который смогли успешно интегрировать в нужный участок генома. Получилось сделать это у них за счет явления, ныне известного как «гомологичная рекомбинация», и далее мы о нем расскажем чуть подробней .
Между прочим, эти опыты легли в основу метода «нокаута» генов, широко используемого в биомедицине и доклинических исследованиях. В 2007 году Смитис вполне заслуженно получил за свои открытия Нобелевскую премию, разделив ее с двумя другими выдающимися исследователями: Марио Капекки и Мартином Эвансом.
Исследования Смитиса были, безусловно, новаторскими: в то время это был очень важный результат для фундаментальной науки; но для науки прикладной пока еще было мало толку. Дело в том, что эффективность генной интеграции тогда была очень низкой: примерно одно событие на тысячу клеток. К тому же, даже в успешно трансформированных клетках продукция целевого белка наблюдалась не всегда, так что до генного редактирования было еще далеко.
Чуть ближе эпоху «генных правок» приблизила группа ученых во главе с Лоиком Перруо из Национального центра научных исследований во Франции. Они синтезировали недлинные нуклеотидные цепочки, способные под воздействием активирующего их света расщеплять нуклеиновые кислоты в определенных местах. Такие вот специфические искусственные фотоэндонуклеазы были своего рода прототипами-предшественниками современных режущих ДНК ферментов — рестриктаз. Последние, правда, ученым уже не пришлось искусственно синтезировать — они «позаимствовали» их из живой природы.
«Отбирая» дары эволюции
Сделаем небольшое (лирическое, но подводящее к главному) отступление. Когда для медицинских целей ученым потребовались обладающие биологической активностью крупные молекулы, они сначала пробовали синтезировать их химическим путем. Так, например, пытались делать инсулин, синтезировав его искусственно последовательным пришиванием по одной аминокислоте в цепь (до победного!)… Производство таким путем не особо «заладилось», что в целом неудивительно: даже для относительно небольшой молекулы инсулина это было чересчур долго, дорого и сложно. Здесь-то на помощь и пришли методы микробиологии и генной инженерии: «приручив» кишечную палочку (читай, генетически модифицировав ее для производства инсулина), исследователи смогли ее культивировать в биореакторах, а продуцируемый ею целевой белок снимать как сливки ко всеобщей пользе .
Во всем этом удивительного тоже мало: фармпромышленность так и развивалась — от производства природных экстрактов и препаратов из них к получаемым методами химического синтеза малым молекулам, а затем к крупным лекарствам-биомолекулам — и далее к передовой терапии. Собственно, производимый биотехнологически инсулин так и появился, а за ним и множество других замечательных биопрепаратов. Обо всех этих «метаморфозах» мы подробно рассказали в статье «Три поколения лекарств» спецпроекта «Современные лекарства».
Этот простой пример показывает, что при выходе на новый уровень молекулярной сложности гораздо проще (а главное, эффективней) не изобретать велосипед, а просто взять его из природы, слегка (а может даже и существенно) модифицировав под нужды науки. В генном редактировании такой подход помог создать по-настоящему эффективные инструменты.
Что было дальше?
Ниже немного расскажем об исследованиях, без которых «генные редакторы» вряд ли бы появились, так что об этом трудно смолчать, ведь «из песни слов не выкинешь». Во всех красках история появления инструментов молекулярной биологии и генной инженерии рассказана в статье «12 методов в картинках: генная инженерия. Часть I, историческая»; здесь же мы обозначаем лишь некоторые ее ключевые эпизоды.
Молекулярные ножницы из недр природы
В 1978 году Нобелевскую премию по физиологии и медицине «за открытие рестрикционных ферментов и их применение к проблемам молекулярной генетики» разделили микробиологи Вернер Арбер, Даниел Натанс и Гамильтон Смит. Каждый из них внес свою лепту: Арбер показал, что кишечные палочки расщепляют ДНК фагов лямбда особыми ферментами-рестриктазами (или эндонуклеазами); Гамильтон такие ферменты выделил уже из гемофильной палочки; ну а Натанс выделил и охарактеризовал фрагменты нарезаемой рестриктазами ДНК, продемонстрировав практическую пользу этих «ножниц».
После всех этих открытий, казалось бы, «карты в руки»! Рестриктазами можно разрезать ДНК, вставляя отрезанное из одного организма в другой: вот они возможности для генной инженерии и модификации! Стало быть, можно уже начинать «кройку и шитье» геномов, исправляя природные недостатки и заболевания?..
Как выяснилось, не совсем. Дело в том, что хотя природные эндонуклеазы разрезают ДНК в предопределенных местах, иногда это совсем не те места, что нужны ученым. Обладая своей собственной сайт-специфичностью, эти ферменты оказались «капризными лошадками», которых не так легко приручить. Вшитое эволюцией нацеливание на расщепление строго определенных последовательностей изменить не так просто, а потому для «свободного» редактирования генома потребовался гораздо более тонко настраиваемый инструментарий. Необходимо было продолжать поиск новых решений, и к счастью ученые здесь не сбавляли обороты.
Мегануклеазы
Важной вехой в преодолении ограничений стало открытие, что если разрезать сразу обе цепи ДНК (сделать двухцепочечный разрыв — double strand break; DSB) в образовавшуюся прореху будет гораздо легче интегрировать требуемый генетический материал.
В этих исследованиях использовали так называемую мегануклеазу I-SceI, полученную из обычных пекарских дрожжей (Saccharomyces cerevisiae). Как выяснилось, она способна разреза́ть обе цепи ДНК в строго определенном месте, причем ее уникальный сайт распознавания состоит из 18 пар нуклеотидов, что практически исключает его случайное присутствие в геноме (в отличие от паттернов расщепления «обычных» рестриктаз, образованных 4–8 парами оснований). Например, геном человека пришлось бы увеличить 20-кратно, чтобы «внезапное» появление в нем сайта мегануклеазы стало возможным. Такая эксклюзивность позволила редактировать гены гораздо точней и эффективней: в частности, производить «правки» внутри организмов (in vivo), а также детальнее изучить последствия двухцепочечных разрывов «молекулы жизни».
Оказалось, что после расщепления обеих нитей ДНК у клетки появляется два пути ее «ремонта»: путем негомологичного соединения концов (грубо говоря «сшиванием») и методом гомологичной рекомбинации. Первый способ чреват ошибками: образованием случайных инсерций и делеций (то есть нуклеотидных вставок и «утрат»); второй же позволяет внедрить в ДНК требуемую последовательность используемого шаблона для репарации, приводя тем самым к желанному результату. Тут не только с большей точностью восстанавливается ДНК, но и нужным исследователям образом меняется ее цепочка (а это, по сути, и есть редактирование).
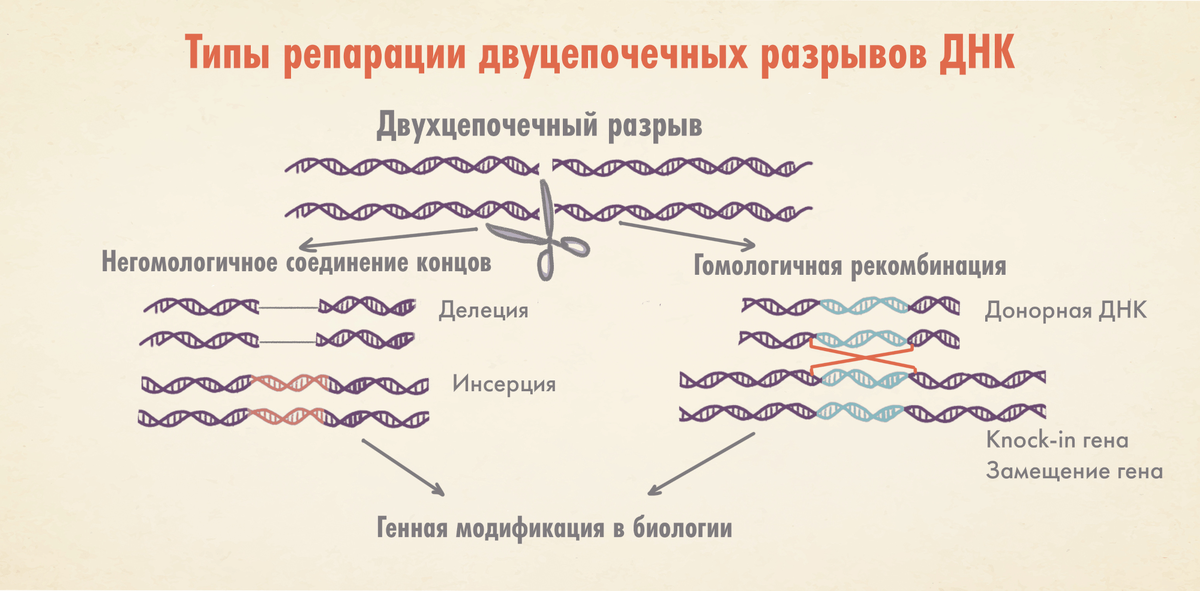
Мегануклеазы, собственно, и стали одними из первых по-настоящему годных инструментов генного редактирования; правда, оказались они совсем не без изъянов. Дело в том, что хотя в природе таких ферментов существуют сотни, их строго фиксированная сайт-специфичность резко ограничивала выбор возможных целевых локусов. Впоследствии сайты распознавания мегануклеаз переделали под более гибкую настройку, позволив проводить направленное редактирование в клетках млекопитающих — мышей и крыс. К сожалению, «перекраивать» мегануклеазы под каждую новую задачу оказалось делом затратным и (на первый взгляд) не особо перспективным. Вот почему их применение в медицине пришлось отложить в долгий ящик, ну а поиск новых инструментов для модификации генома тем временем продолжился.
Сейчас уже есть пример их использования в медицине, расскажем о нем ниже.
Нуклеазы с цинковыми пальцами (Zinc-finger nucleases — ZFN)
Еще в 1980-х ученые выделили из бактерий Flavobacterium okeanokoites эндонуклеазу Fok1, способную к неспецифическому разрезанию ДНК благодаря наличию особого каталитического домена. Фермент этот, казалось бы, универсален: бери и режь им геном в любых местах; но в том-то и дело, что для целей редактирования специфичности ему тогда как раз и не хватало…
Дабы наделить Fok1 «нацеливающими» свойствами, ученые соединили ее каталитический домен с так называемыми цинковыми пальцами — ДНК-связывающими фрагментами транскрипционных факторов.
Благодаря своей «природной миссии» каждый «палец» строго комплементарен определенному триплету, что уже задает специфичность. Эта природная особенность позволила настраивать инструменты геномного редактирования под нужды исследователей: варьируя тип и количество цинковых пальцев достигать требуемого «нацеливания». То есть в теории, имея «пальцы» для каждого из 64 возможных кодонов, можно собирать белковые модули, направленные на любую желаемую последовательность ДНК, что позволяет разреза́ть геном где угодно.
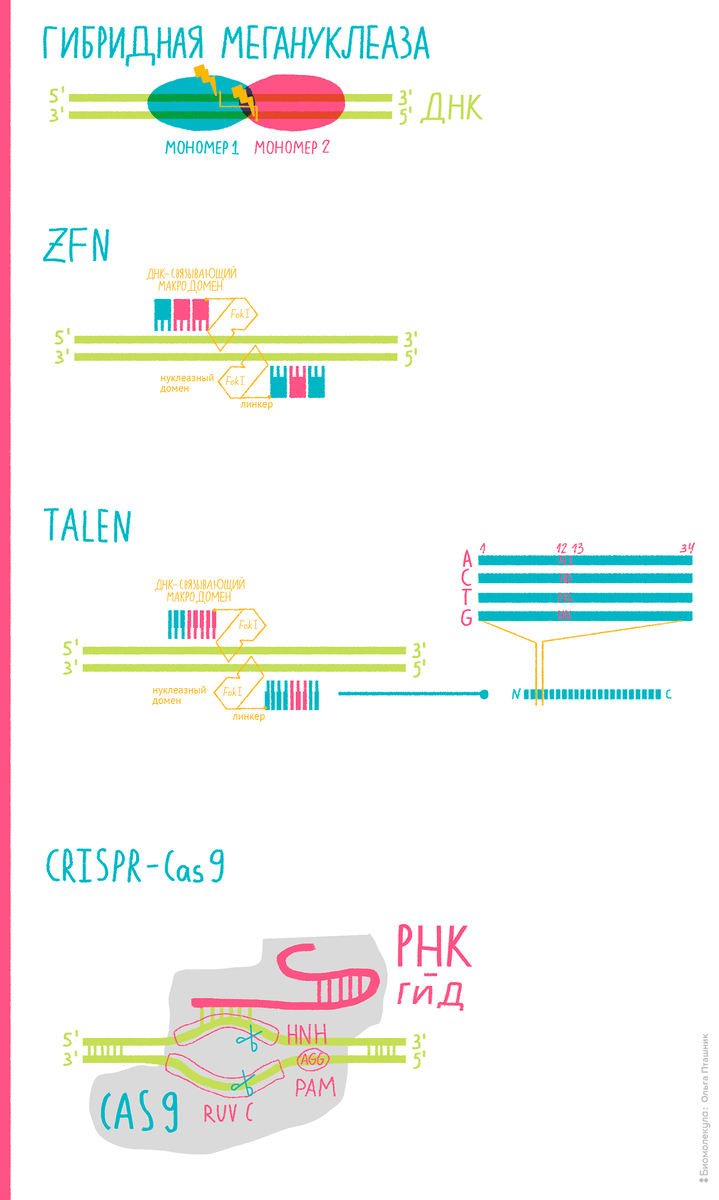
Так вот, связав структурные возможности цинковых пальцев с «режущими способностями» Fok1, ученые получили прекрасный модульный инструмент генного редактирования, пригодный под самые разные нужды (рис. 2). Когда его разработали, им не преминули воспользоваться для редактирования геномов различных организмов: мух Drosophila, нематод C. elegans, морских ежей, тутовых шелкопрядов, рыбок Danio rerio, шпорцевых лягушек, мышей, крыс, кроликов, свиней, коров и др.; но что самое важное — ZFN использовали при геномной модификации различных человеческих клеточных линий [13]. Этот этап редактирования клеток человека, запущенный в 2005 году нашим выдающимся соотечественником Федором Урновым, стал началом сложного пути, который в конечном итоге и привел к лечению наследственных заболеваний с помощью инструментов редактирования.
Впрочем, для полноценного использования у человека технологии генных манипуляций еще требовали значительного улучшения.
Эффекторные белки, подобные активаторам транскрипции
Следующим улучшением стало объединение «режущих» нуклеаз Fok1 со специфичностью белков TALE, выделенных из бактерий рода Xanthomonas. Эти белки, как и следует из их названия, подобны активаторам транскрипции (TALE = Transcription activator-like effectors). Xanthomonas впрыскивают их в растительные клетки, и там уже эти ферменты осуществляют совершенно особое действо: «садясь» на ДНК клеток-хозяев, начинают модулировать экспрессию их белков выгодным для этих бактериальных паразитов образом.
Чтобы вот так ловко перепрограммировать генетические механизмы белкового синтеза растений, ксантомонады наделили свое «оружие» TALE выдающимися свойствами. Каждый их тандемный повтор строго специфичен для определенного нуклеотида — это-то и сделало их использование в геномном редактировании привлекательным для исследователей. В паре с нуклеазами Fok1 они обеспечивают бо́льшую точность редактирования в сравнении с ZFN.
Недостающее звено
При всех плюсах TALEN и ZFN, существенный их минус — в недостаточной воспроизводимости и гибкости. Ведь для каждого нацеливания на новый геномный локус эти инструменты потребуют повторения сборки белковых модулей с заданной специфичностью, что занимает много времени, непрактично и обходится совсем недешево .
Что-то подобное можно сказать и про мегануклеазы, ведь их специфичность «настраивается» генной инженерией; о других недостатках и преимуществах различных систем редактирования см. предыдущую статью, а также материал «12 методов в картинках: генная инженерия. Часть II: инструменты и техники» (там этот вопрос разобрали несколько подробней).
Прорыв, сделавший геномное редактирование гораздо более практичным и широкодоступным, связан с разработкой знаменитой системы CRISPR-Cas9, о которой на «Биомолекулe» писали много и со вкусом, что, кстати, неудивительно: метод этот со временем обрел заслуженную славу и популярность среди исследователей и медиков (ниже рассказываем, почему).
А что потом? О CRISPR-Cas9 и еще больше о генном редактировании и его использовании в современной медицине вы узнаете из нашей новой статьи!