Вступление
Цинодонты возникли в межкризисную эпоху лопингского века (поздней перми), сразу же после массового вымирания под конец каптанского яруса. В поздней перми цинодонты ещё не успели проявить себя в должной мере, а потому сидели в тени более крупных и сильных сородичей – сначала горгонопсид, а затем и тероцефалов которые пришли на замену первым когда климат стал ухудшаться.
Впрочем нельзя сказать что они не пытались, южноафриканский цинодонт ветусодон (Vetusodon) был одним из главных хищников в своей экосистеме, уступая лишь более крупному тероцефалу мосхорину (Moschorhynus). Но продлился век ветусодона недолго и уже совсем скоро на Землю с огромной силой обрушилось новое разрушительное вымирание унёсшее 95% всей жизни на Земле, забрав в мир иной в том числе и ветусодона.
Однако другим, более мелким цинодонтам, таким как тринаксодон, повезло больше и они смогли пережить вымирание смело перейдя в новую, мезозойскую эпоху. Но и в постапокалиптическом раннем триасе цинодонты ещё не спешили идти на вершину пищевой цепочки, оставив ниши растительноядных дицинодонтам, а ниши хищников архозаврообразным таким как протерозух и эритрозух.
Цинодонты решили рвануть в крупноразмерные ниши хищников только под конец оленёкского яруса ~247,2 миллиона лет назад. К крупнейшим цинодонтам жившим тогда можно отнести всеядного диадемодона (Diademodon tetragonus) и гиперплотоядного Циногната (Cynognathus), про которого будет поведано далее.

Краткий очерк
Циногнат, или даже киногнат (лат. Cynognathus, название происходит от древнегреческого словосочетания κύνεος γνάθος и переводится как "собачья челюсть", или "клыкастая челюсть") – вымерший род цинодонтов из клады циногнатий, живший на территории Южной Африки, Антарктиды, Аргентины, Намибии и возможно Китая. Типовой и единственный вид внутри рода Cynognathus crateronotus.
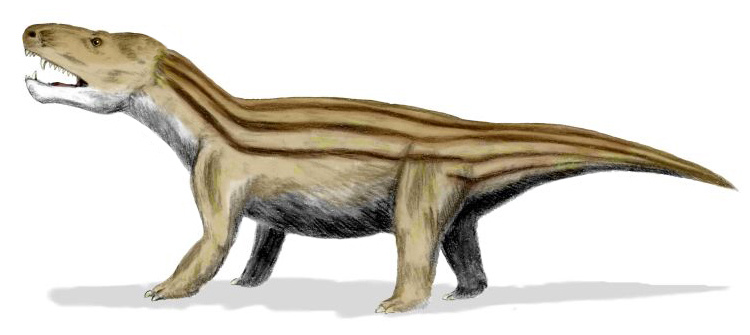
Род циногнатов существовал достаточно продолжительное время в среднем триасе с начала анизийского по конец ладинского яруса 247,2(возможно и 249)-237/236 миллионов лет назад, т.е. в общей сумме 10+ миллионов лет, что весьма и весьма похвально.
Род находится основании одноимённой клады – циногнатии (Cynognathia), в которую входят так же 3 семейства других цинодонтов. Помимо этого циногнат образует своё собственное семейство – циногнатиды.
Описание
Размеры
Циногнат был крупным цинодонтом достигавшим длины в 1,2 метра без хвоста и 2 метра с хвостом. Длина черепа циногната достигала 40 см.
Но...
В статье про ранний триас было написано о том что циногнат мог спокойно вымахивать до 3 метров в длину и веса в 300+ кило. Это частично верная информация ведь в Намибии был найден неописанный экземпляр гигантского циногната с длиной черепа 70 сантиметров!
Череп
Череп большой относительно остального тела. Его предглазничный отдел вытянутый, сужающийся позади крупных клыков. Глазница небольшая. Заглазничная дуга широкая. Височное окно очень велико и подымается до теменной кости, заглазничная и чешуйчатая кости уже не соприкасаются друг с другом над этим окном. Окологлазничный ряд костей редуцировался; предлобная и слезная кости малы, а заднелобная исчезла. Скуловая дуга массивная и широко выступает наружу; ее верхний край расположен много ниже сагиттального гребня. Сзади скуловая дуга широко соединяется с мозговой коробкой. Аддукторная камера расширена наружу и назад; окружающие ее кости шире и толще, чем у тринаксодона, что свидетельствует об очень сильном укусе. Камера не тянется так далеко вперед, значит, челюсти раскрывались шире, позволяя схватить более крупную добычу.
Предтеменная кость отсутствует. Пинеальное отверстие очень мало или отсутствует. Сагиттальный гребень хорошо развит. Квадратноскуловая кость мала и тесно прирастает ко столь же редуцированной квадратной; обе эти кости довольно слабо соединены с остальным черепом. На затылочной поверхности чешуйчатой кости имеется отчетливый желобок, который, как полагают, является наружным слуховым проходом, этот желобок ведет сверху вниз к челюстному суставу. Затылок широкий, возможно, для прикрепления очень массивной туловищной мускулатуры; латерально он сливается с концами очень высоких скуловых дуг. Затылочная поверхность черепа является почти сплошной пластинкой, состоящей из соединенных затылочных и верхнеушных костей вместе с лежащими выше кожным верхнезатылочными и табличатыми костями. Затылочный мыщелок разделился и представляет собой двойное образование.
Продолжениями предчелюстных, верхнечелюстных и небных костей образована пластинка, которая лежит горизонтально ниже прежнего уровня крыши рта, так что образуется полное вторичное небо. Хоаны перемещаются назад через пространство, находящееся над вторичным небом. Оба предсошника срастаются в единое образование, от которого по срединной линии отходит отросток. Небные зубы отсутствуют. Межптеригоидных ям тоже уже нет, поскольку оба птеригоида срослись друг с другом в одно сплошное образование под передним концом черепной коробки; парасфеноид сохраняется в своем старом положении выше них и между ними. Птеригоиды укоротились и больше уже не протягиваются назад к квадратным костям, хотя расположенные выше них эпиптеригоиды могут продолжаться назад до этих последних. Сохраняются уменьшенные поперечные отростки птеригоидов. В дополнение к обычным костям черепной коробки эпиптеригоид образует плоскую пластинку, примыкающую к черепной коробке впереди переднеушной кости, а в более передней части имеется, по-видимому, пара костей, соответствующая орбитосфеноидам млекопитающих.
Зубная кость составляет значительно большую часть нижней челюсти, чем у более примитивных цинодонтов. Внешне она напоминает таковую крупных хищных млекопитающих. Назад и вверх от зубной кости отходит массивный венечный отросток с выпуклым задним краем, почти касающимся чешуйчатой кости. Яма жевательной мышцы глубокая, заходит далеко вперед. Нижний край зубной кости продолжается назад угловым отростком. Нижне-задний угол зубной кости выступает назад от края нижней челюсти. Задние кости нижней челюсти значительно редуцированы и объединены в узкий стержень, вставленный в желобок на медиальной поверхности зубной кости. Он состоит из сочленовной, предсочленовной, угловой и надугловой костей. Венечная кость остается в виде плоской пластины, прикрывающей передний конец этого стержня. Тип челюстного состава такой же, как у примитивных терапсид; он образован сочленовной и квадратной костями. Стремя тянется в виде тонкой палочки от отверстия внутреннего уха к квадратной кости.
Ямы и каналы на лицевой поверхности черепа указывают на концентрации нервов и кровеносных сосудов, которые могли быть связаны с вибриссами.
Зубы отчетливо дифференцированы, их формула: I4/i3, C1/c1, Pc9-10/pc9. Резцы и крупные клыки конические, несколько загнутые назад. Заклыковые зубы сжатые латерально, грубо пильчатые. Большинство задних заклыковых зубов трехзубчатые, с более мощными, чем у Thrinaxodon, зубцами и отклоненной назад главной вершиной, но на самых задних зубах появляются дополнительные вершины, расположенные позади главной. Лингвальный поясок на заклыковых зубах не развит. Обычно его отсутствие считают вторичным явлением, связанным с питанием циногната более крупной добычей. Предпоследний зуб увеличен и аналогичен хищническому зубу плотоядных млекопитающих. Наблюдается некоторое истирание зубов, но точного прикуса между верхними и нижними зубными рядами нет.
Посткраниал
Телосложение довольно массивное. Туловище вытянутое. В позвоночнике исчезли интерцентры, хотя тела позвонков остаются амфицельными. Насчитывается 6-7 шейных, 17-18 грудных, 4-5 поясничных и 3-4 крестцовых позвонка. Свободно подвижные двуглавые ребра имеются на всем протяжении от шеи до основания хвоста, постепенно укорачиваясь кзади. На задних спинных ребрах сохраняются расширенные пластинки, характерные для тринаксодонов. Сохраняются гастралии.
Плечевой пояс продолжает состоять из кожного пояса, в который входят межключица и ключицы (клейтрум исчез), и первичного пояса из лопатки и 2 коракоидных элементов. Передний край лопатки отвернут кнаружи (начало возникновения гребня, свойственного млекопитающим), и ключица прикрепляется к отростку, выступающему у нижнего конца гребня. В тазовом поясе подвздошная кость вытягивается скорее вперед, чем назад (последнее положение является примитивным), в связи с изменением положения мышц, и прикрепляется к возросшему числу крестцовых ребер. Лобковая и укороченная седалищная кости передвинулись назад (что также связано с изменением в положении мышц); между ними появилось хорошо развитое запирательное отверстие, которое частично разделяет их.
Передние конечности несколько расставлены в стороны, а задние размещаюися непосредственно под телом. На бедренной кости головка развивается скорее сбоку, чем на конце этой кости, в связи с тем, что она повернута кпереди; а на наружной стороне имеется большой трохантер для мускулов, которые идут от подвздошной кости. Пальцы, за исключением I-го, имеют почти одинаковую длину в связи с тем, что конечностям свойственно передне-заднее движение. Фаланговая формула составляет 2 - 3 - 4 - 5 - 3, так как в III-м и в IV-м пальце сохраняются рудименты лишних фаланг.
Кортекс длинных костей конечностей выраженно зонален, построен из фиброламеллярной ткани и богато васкуляризирован, тогда как ламеллярная ткань в нем отсутствует. Это свидетельствует о непрерывности роста костной ткани.
Палеобиология и палеоэкология
Палеогеография
Циногнат был типичным компонентом среднетриасовой фауны захватив многие экосистемы в Африке (бассейн Кару, группа Бофорт), а точнее в Намибии, Южной Африке и Лесото, где специально выделяется зона скопления Циногнатов. Помимо этого останки циногната известны из формации Фремоу в Трансантарктических горах Антарктиды и формации Пуэсто-Вьехо в Аргентине.
По сути циногнат сумел частично повторить успех листрозавра, который так же был распространен во многих экосистемах своего времени.
Образ жизни
Диета
Циногнат был хищным животным который питался как мелкой так и крупной добычей, а также не брезговал падалью.
В поедании пищи ему сильно помогали его зубы а также вторичное нёбо, позволявшее глотать пищу и дышать одновременно.
Стайность или одиночество?
Неизвестно был ли циногнат стайным животным или собирался в группы только для охоты на крупную добычу.
Одна из гипотез говорит о том что циногнаты во многом были успешны благодаря групповой охоте как современные волки, тогда как одиночные терапсидные охотники такие как мосхорин был вытеснены (ну или заменён) архозаврообразными протерозухами во многом из-за образа жизни аналогичного современным кошачьим.
Однако эта гипотеза имеет мало подтверждений и всё таки циногнат является одиночным хищником.
Добыча и палеоокружение
Во времена циногната главными растительноядными животными всё ещё оставались дицинодонты, в частности процветавшее в то время семейство каннемейериид. Циногнат пересекался и охотился на таких дицинодонтов как: собственно Каннемейерия, ангонизавр, долихуран, тетрагоний, сангузавр, замбиязавр и т.д.
Помимо этого в рацион циногната могли входить животные и поменьше такие как мезозух, эупаркерия, стенавлоринх а также темноспондильные амфибии потипу антарктозуха. Добычей мог стать и соразмерный циногнату его близкий родич – диадемодон.
Конкуренцию циногнату составляли самые разные хищники, большинство из которых он успешно пережил, среди них есть такие гиганты как архозаврообразный эритрозух, крупный цинодонт трираходонтид импиденс и близкий родич динозавров телеократер.
Палеосреда
В бассейне реки Кару на территории современной Южной Африки циногнат обитал в открытой местности с тёплым сухим летом и прохладной влажной зимой, похожей на современный регион Западный Кейп, но более засушливой. Особи росли быстро и непрерывно. Смена времён года практически не влияла на их охотничьи привычки.