Введение
Клада цинодонтов возникла во время межкризисной эпохи под названием поздняя пермь или же Лопингский/Татарский век ~259,59 миллионов лет назад. В то время это были маленькие животные, чаще всего довольствовавшиеся насекомыми, правда были и те кто по образу жизни напоминал выдру, как например процинозух. Отдельные представители клады даже смогли пробиться в ранг среднеразмерных хищников как например южноафриканский ветусодон. Впрочем если же говорить про абсолютное большинство то оно всё же оставалось в тени своих териодонтных родичей – горгонопсид и тероцефалов.
Так бы дела дальше и шли, если бы на горизонте не замаячило одно событие, координально поменявшее образ жизни данных существ, – великое пермское вымирание произошедшее 251,9 миллионов лет назад и унёсшее жизни 95% существ на Земле, как в воде так и на суше. Цинодонтам тоже досталось, впрочем для них это вымирание было не таким катастрофическим как для их сородичей (и возможно предков) тероцефалов, весь цвет которых был отправлен в мир иной.
Впрочем цинодонты что до что сразу после вымирания вставать на ноги не спешили, они всё так же оставались в нише мелкоразмерных хищников аки тринаксодон или трираходон из раннего триаса. Однако время благоволило нашем родичам ведь после вымирания многие экосистемы опустели, а потому настало наиболее подходящее время для увеличения разнообразия и попутного освоения новых ниш.Конкуренции было мало, а потому оказать достойный отпор цинодонтиям никто не мог, поэтому уже к началу среднего триаса цинодонты были уже во все оружия и рвались к покорениям новых высот.
Особо на этом фоне выделилась группа эпицинодонтов-циногнатий – продвинутых цинодонтов нового типа, которая породила таких чудищ как циногнат, импиденс или как например герой данной статьи, – Диадемодон, о нём собственно и будет подробно рассказано здесь.
Краткий очерк
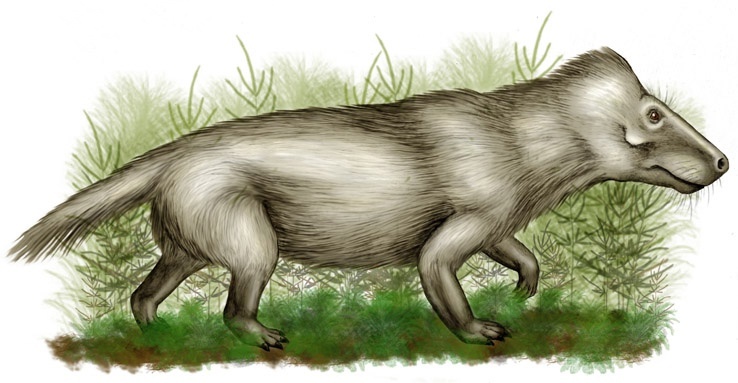
Диадемодон (лат. Diademodon) – вымерший род гомфодонтных цинодонтов живший на территории Гондваны, в частности ЮАР, Замбии, Намибии, Зимбабве, Танзании и Аргентины (формация Рио-Секо-де-ла-Куэбрада, Мендоса) в анизийском ярусе среднего триаса ~247-237 миллионов лет назад. Типовой и единственный вид Diademodon tetragonus. Диадемодон является типовым родом своего семейства, но не единственным его представителем, там же рядышком уместился его близкий родич титаногомфодон, живший примерно в то же время что и сам диадемодон.

Описание
Размеры
Диадемодон был достаточно крупным цинодонтом достигавшим от 1,5 до 2 метров в длину. Длина черепа была 20-30 сантиметров.
Анатомия
Морда довольно узкая и, как у циногната, еще сильнее сужается позади клыков. Заглазничная дуга относительно узкая. Скуловая дуга очень высокая, с вырезкой у переднего основания, отделяющей ее от максиллы. На переднем конце скуловой дуги имеется массивный отросток для прикрепления жевательной мышцы, отсутствующий у хищных цинодонтов. Как и у циногната, но в отличие от тринаксодона, задний конец чешуйчатой кости выдается латерально, образуя желобок, идущий вниз до области челюстного сочленения (это образование очень характерно для всех гомфодонтов, но менее выражено у других цинодонтов). Аддукторная камера гораздо крупнее, чем у циногната. Сохраняются маленькое пинеальное отверстие и поперечные отростки птеригоидов. Имеется 2 раздельных затылочных мыщелка.
Венечный отросток зубной кости нижней челюсти очень высокий и широкий, но, в отличие от циногнатид, вогнутый по заднему краю. Ретроартикулярный отросток сочленовной кости слегка изогнут назад. Угловой отросток зачаточный, но выражен лучше, чем у циногнатуса. Постдентальные кости сильно редуцированы, при этом угловая кость уменьшается до узкого стержня, параллельного предсочленовной и надугловой костям. Как и у других терапсид, сочленовная и квадратная кости являются элементами челюстного сустава, но соединяются они друг с другом точно таким же образом, как у млекопитающих сочленяются гомологи этих костей — молоточек и наковальня, а квадратная кость соединяется со стременем.
В верхней челюсти имеются 4 резца, клык и до 16 заклыковых зубов, в нижней — 3 резца, клык и до 12 заклыковых зубов. За большими верхним и нижним клыками есть небольшие диастемы. Вверху молодые особи имеют всего 4 "предкоренных" зуба, у взрослых общее число заклыковых зубов возрастает до 16. Передние 4-5 заклыковых зуба простые конические, следующие за ними — латерально сжатые и режущие, как у примитивных цинодонтов. В отличие от циногнатов, задние заклыковые зубы поперечно расширены и обладают относительно плоскими вершинами — в результате этого они способны давить и перетирать пищу. Величина заклыковых зубов увеличивается кзади, но последние 3 зуба вновь уменьшаются. Верхние зубы значительно шире, чем нижние.
Задние, расширенные заклыковые зубы снабжены бугорками: на больших зубах — по 4 или 5, последний, маленький, имеет 2 бугра: один — большой, загнутый в виде конуса, другой — маленький, придаточный. После прорезания зубов эмаль образовывала ряд мелких бугорков по периферии коронки и крупный бугор у середины наружного края, соединенный со слабо выраженным поперечным гребнем. По мере износа эмаль на вершине зуба утрачивалась, оставаясь лишь вокруг дентинового цилиндра. На протяжении жизни происходила замена резцов, клыков и 4 передних заклыковых зубов.
В поясничной области сохраняются широкие реберные пластины, как у тринаксодона, однако на шейных и передних грудных ребрах такие пластины отсутствуют. В отличие от циногната, конечности относительно короткие. Кортекс длинных костей конечностей выражено зонален, построен из чередующихся колец фиброламеллярной и ламеллярной тканей. Это свидетельствует о том, что рост костной ткани происходил циклически, с перерывами.
Образ жизни
Диадемодон был достаточно распространённым животным в экосистемах Гондваны. Согласно тафономическим данным, он населял преимущественно полупустынные местности с резким разделением года на сухой и дождливый сезоны. Полагают, что диадемодон был всеядным, питаясь как другими животными и падалью, так и мягкой растительностью.
Некоторые предположения и вовсе говорят о полуводном образе жизни диадемодона, а-ля эдакого бегемота который питался водорослями и днём оставался в глубоких водоёмах, а ночью покидал водную среду, чтобы добывать пищу. Это было установлено по низким показателям содержания кислорода в образцах тканей. Но данная идея была опровергнута, поскольку диадемодон не имел морфологических или изотопных признаков адаптации к длительному пребыванию в воде. Уровень кислорода в его костях не соответствовал уровню кислорода в костях других полуводных организмов, таких как бегемот. Третья предполагаемая возможность заключалась в том, что диадемодон мог находиться рядом с легкодоступными источниками воды, но не проводил в них большую часть своей жизни. Возможно, он питался мелководными водорослями, в которых, как известно, меньше кислорода.