
ФАКТ 8
Возникнув при становлении сравнительной этологии между 1911 и 1937 гг., «инстинкт» заимствовался в прочие поведенческие дисциплины[34]. Рождение этологии обусловилось гениальным прозрением Оскара Хейнрота, вдруг «увидевшего», что наследственная координация телодвижений и действий животного, стоящий над нею центр торможения и специфические формы стимуляции, запускающие инстинктивный акт «образуют с самого начала некоторое функциональное целое»[35].
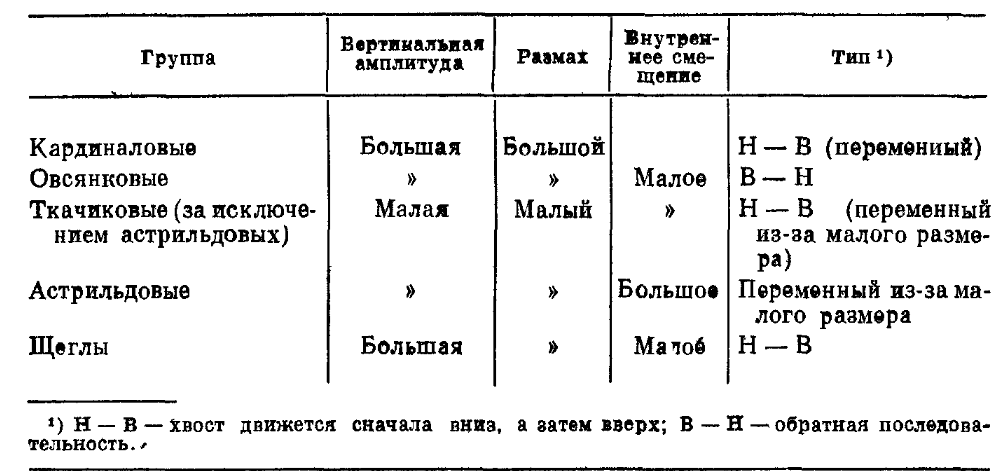
Выделив эту систему, Хейнрот ввёл понятие «присущего виду импульсивного поведения» (arteigene Triebhandlung), расчистившее путь «морфологическому методу» этологов. Arteigene Triebhandlung — та самая «манера держаться», по которой орнитолог уверенно распознаёт вид ещё до того, как рассмотрит детали окраски. Пример: реакции встряхивания хвостом, характерные движения при взлёте, чистке и пр., другие аспекты манеры держаться у разных групп птиц настолько устойчивы и типизированы, что имеют систематическое значение [36] (рис.8).
Тем более это верно для форм демонстраций, выделенных морфологическим методом: их можно использовать для нужд таксономии и исследований эволюции не хуже «обычных» морфоструктур, а часто и лучше. Прежде всего, дифференциация форм гомологических демонстраций в разных популяциях отражает начало процесса видообразования. Так, у самцов классического объекта этологов — трёхиглой колюшки Gasterosteus aculeatus этограмма ухаживания почти неизменна в широком голарктическом ареале. Однако количественно стереотип полового поведения и особенно динамика его «диалогового» взаимодействия с самкой сильно разнятся между популяциями (собственно поведение самок у всех колюшковых не является видоспецифичным).
Нерест колюшек чётко делится на подготовительную и нерестовую фазы. В первой побуждения самца в присутствии самки перестраиваются: падает агрессивность, растёт половая мотивация. Ей также присуще резкое снижение частоты/интенсивности обмена стимулами между партнёрами, появление вместо них смещённых или переадресованных действий самца, не соответствующих контексту. Смещения связаны со строительством или вентилированием гнезда. Нерестовая фаза — прямое привлечение самки к гнезду и вымётывание икры.
В разных географических популяциях сильней всего различается именно подготовительная фаза стереотипа ухаживания: варьирует длительность, частота и ритуализованность элементов, прежде всего зигзагообразных танцев, а также порядок их следования в составе общевидового ритуала. Беломорские колюшки демонстрируют два разных типа «подталкивания» самцом входящей в гнездо самки. У 30% из них оно не отличается от «подталкивания» колюшек балтийской и черноморской популяций, но 70% самцов «подталкивают» самок по типу, присущему рыбам из Японского моря. Самцы последних практически лишены знаменитого «зигзагообразного танца», типичного для европейских и американских колюшек. Точней, движение, производимое самцом на «нужном» этапе развития взаимодействия, так слабо ритуализовано, что специфический «зигзаг» почти не распознаётся.
Нерестовая фаза, в общем, консервативна и не показывает сколько-нибудь заметных межпопуляционных различий. Вспомним, что лишь в начале ухаживания самка имеет определённую свободу выбора ответа на демонстрации самца. В нерестовой фазе рыбка направляется к гнезду воздействиями от самца, пусть даже стимуляцией, а не насилием, в этих условиях она лишь отказаться от нереста, уплыть прочь, но не вариировать свою реакцию при его продолжении[37]. Поэтому зафиксированная дифференциация брачных сигналов в разных частях ареала G.aculeatus маркирует начавшуюся дивергенцию соответствующих коммуникативных сообществ а, значит, и популяционных систем, давая в потенции новые подвиды и виды.
Так, о. Гаити населяют три аллопатрических вида анолисов, почти неразличимые морфологически – Anolis brevirostris, A. websteri и A.caudalis. Однако они чётко различны по демонстрациям ухаживания-угрозы между самцами, конкурирующими за самку. Анализ 736 отснятых на киноплёнку серий подобного поведения всех видов (n=46 самцов) показал, что их демонстрации в целом гомологичны, но различаются вариантами исполнения одного и того же комплекса фиксированных действий. Одна из альтернатив исполнения регистрируется у одного вида, другая у другого, и некоторая часть демонстраций тождественна у всех трёх (Jenssen, Gladson, 1984).
Тип демонстраций А характеризуется серией из 5 «глубоких» кивков головы с 1-2 добавочными, без раздувания горлового мешка, В — много бόльшим (и нефиксированным) числом «мелких» кивков, горловой мешок расправлен и пульсирует. A.caudalis обладает одним вариантом В, A.brevirostris — двумя дифференцированными вариантами, чётко отличными от той же позы A.caudalis, A.websteri — тремя. По впечатлениям наблюдателя-этолога А неразличимы у всех трёх, однако дискриминантный анализ устойчиво разделяет их по промерам реализаций данных поз, сделанных по кадрам киносъёмки. Различия сигнальных репертуаров чётче всего выражены между географически близкими первым и третьим видом; авторы объясняют это «смещением признаков» (Jenssen, Gladson, 1984), но, судя по всему, это просто следствие разной длины филогенетической дистанции.
За актами видообразования с дивергенцией видов как «замкнутых» популяционных систем следует дивергенция коммуникативных сообществ, присущих им в меру «социальности» вида. Это вызовет столь же закономерную дивергенцию форм демонстраций, обслуживающих взаимодействия животных в сообществе, обеспечивающих коммуникацию в обоих системах, материнской и дочерней. В этом случае степень несходства формы сигналов (сумма различий, накопленных в соответствующих репертуарах из m сигналов) у i дивергировавших форм в некоторой филогенетической ветви примерно пропорциональна времени дивергенции.
ФАКТ 9
Если типизированность и узнаваемость облика присущи даже повседневной активности (не континуальной, но структурированной благодаря Arteigende Triebhandlungen), то тем более это верно для инстинктивного поведения, как релизеров, так и «высвобождённых» ими инстинктивных ответов. Те и другие не «размазаны» в потоке активности, а представляют собой дискретные единицы, узнаваемые благодаря характерной форме. Этологи их зовут «демонстрациями», обычно с добавлением «ритуализованные» (англ. ritualised demonstrations, ritualised postures): они могут быть визуальными (позы), акустическими, запаховыми и следовыми метками, электрическими, даже вибрационными.
Эти отдельности — единицы видового репертуара, дискретизированные в отслеживаемом потоке активности, если (или когда) их специфический облик характеризует именно данную единицу, а не контекст, и не категоризирующее впечатление наблюдателя (управляемое потребностью в простоте и удобстве классификации наблюдаемого). Что видимые им демонстрации именно таковы — это гипотеза, которая в каждом отдельном случае должна быть проверена и может быть опровергнута. Вопреки усилиям критиков сравнительной этологии сегодня точно известно, что многие демонстрации именно таковы.
Больше того, они коммуникативно действенны (как релизеры и/или знаки семиотической системы вида) именно и только тогда когда характеризуются специфическим инвариантом формы. И наоборот — сохранение этой инвариантности облика демонстратором вопреки разного рода помехам, по большей части рождаемым и усиливающимся в самом взаимодействии, однозначно свидетельствует в пользу сигнальности и знаковости соответствующих единиц.
Независимо от способа продуцирования и воспринимающего анализатора, всем демонстрациям присуща типологическая определённость формы или что то же самое, инвариантность образов, «вычерченных» комплексом телодвижений и действий, образующим каждую подобную единицу. Скажем, два симпатрических вида-двойника крупных поморников, южнополярный C.maccormicki и бурый C.lonnbergi крайне похожи, даже для специалистов, а демонстрации их высоко гомологичны, включая свойственный чайковым «долгий крик[38]» (англ. Long call).
Особи обоих видов варьируют в уровне возбуждения, в том числе в ситуациях, связанных с исполнением долгого крика: последнее сильно (и одинаково) варьирует у обоих видов, особенно амплитуда и фиксированность отдельных движений[39] (рис.9А, Б).
Однако Pamela J. Pietz (1985) показала, что эта изменчивость не только не «смазывает» межвидовых различий форм долгого крика. Даже наоборот, она ещё больше «подчёркивает» устойчивость их сохранения в ситуации демонстрирования (рис.9В), вопреки переменчивости обстоятельств [40] контекста внутри (уровни мотивации, общего возбуждения и пр.) и вовне особи (какое-то беспокойство, риск нападения хищника, силовое противодействие других птиц или просто их приближение с демонстрациями), При значительном сходстве общего облика «долгих криков» (виды-то близки, и демонстрации гомологичны — также и даже более морфологии), в каждой «точке» образующих их телодвижений С.maccormicki отклоняет корпус назад и вверх много дальше, чем С.lohnbrergi (рис.9В).
А.
Б.
В.
Cтереотипность реализации «долгих криков» у обоих поморников вовсе не нивелирует различия в исполнении, созданные индивидуальными особенностями или обстоятельствами контекста, это попросту невозможно. Однако она сохраняет и даже подчёркивает инвариантность дифференцирующих характеристик долгих криков обоих, несмотря на высокую гомологию. Это же верно для особенностей телодвижений и/или их исполнения, дифференцирующих разные демонстрации ухаживания, угрозы или иного контекста общения в репертуаре одного и того же вида.
Так, демонстрации ухаживания «dip-shaking» и «bob-shaking» западной поганки Aechomorphus occidentalis похожи: сложены гомологичными телодвижениями, их действия развёртываются в одном направлении т.д. Различия только в порядке следования элементов в обоих случаях, и в части движений, появляющихся в ключевые моменты начала и конца исполнения. «Dip-shaking» включает демонстративное плескание водой в начале и позу с низкой шеей в конце демонстрации, «bob-shaking» — нет. Этого достаточно для создания и «подчёркивания» отдельности данных единиц поведения ухаживания (рис.10), поскольку различия устойчиво сохраняются в максимально гетерогенной выборке независимых предъявлений данной демонстрации, а не «смазываются» ростом различий контекста их предъявления, образующем данную гетерогенность (Nuechterlein, Storer, 1982).
Данные нейробиологии также показывают, что добавление уместного (оптимального) уровня шума усиливает сигнал, облегчает его выделение, вместо ожидаемого «смазывания». Сигналы воспринимаются лучше всего при некотором оптимальном уровне «шума» в канале связи, исторически типичном для контекста общения, в котором происходят «обслуживаемые» ими взаимодействия. Это так называемый «стохастический резонанс»: при меньшем уровне «шума» нейроны гораздо слабей реагируют на стимуляцию вообще, в том числе приходящую от других нейронов ансамбля, работой которого сигналы распознаются как таковые и отреагируются выбором «должных» ответов; при большем уровне «шума» они много хуже выделяют из него искомый сигнал.
Дело в том, что взаимодействие, как и нейроны в мозгах участников — системы с нелинейным развитием активности, управляемые положительной обратной связью между демонстрациями, где «шум» упорядочивает активность (Mitaim, Kosko, 1998). Так, анализ критериев выбора самкой демонстрирующего самца показывает отсутствие прямой связи между изменением характеристик самцового образа и выбором самки, если нет помех. Стоит их ввести, связь появляется (Björklund,2002). Благодаря стохастическому резонансу, подавая в нелинейных системах (каковы все нейроны мозга) шум вместе со слабым сигналом, мы усиливаем сигнал, увеличивая его долю в пропорции сигнал-шум (Смарт, 2017: 74-76).
Примечания:
[34] Поэтому отцы-основатели определили сравнительную этологию как «исследование инстинктов» (Tinbergen N. The study of Instinct. Oxford: Clarendon Press, 1951. 228 pp.) или «объективистская теория инстинкта» (Lorenz K. The Оbjectivistic Theory of Instinсt. Paris: Masson, 1956. Р.51-76). Увы, они забыли сказать, что другая сторона той же медали (в смысле, реализации инстинктивного акта) это организованный обмен сигналами во взаимодействиях между компаньонами, т. е. коммуникация животных в сообществе, поддерживающая устойчивую структуру отношений (=социальную структуру), обеспечивающая дифференциацию ролей, установление асимметрии рангов/статусов между исходно «равнокачественными» участниками взаимодействий (хотя и исследовали данный аспект). Здесь очень к месту метафора лингвиста А.Н.Барулина («Основания семиотики». М., 2002. Т.1.): инстинкт есть сложный рефлекс, дуга которого замыкается между особями в рамках социальной коммуникации (не «внутри» неё, как при обычном рефлексе).
[35] Лоренц К. Оборотная сторона зеркала. СПб.: Республика, 1998. С.341.
[36] Показательно, что адаптивного смысла для них до сих пор не нашли, хотя интенсивно искали. Скажем, несостоятельны адаптационистские ответы на вопрос, «зачем трясогузки трясут гузкой» и почему «трясение» организовано так, что его паттерн дискретный и узнаваемый (а не изменчивый сообразно обстоятельствам): 1) трясогузки так вспугивают насекомых из травы, чтобы легче было схватить (напротив, во время кормления они прекращают трясти), 2) Это сигнал о социальном статусе птиц, 3) трясение скоординировано с осматриванием местности, поэтому может быть сигналом о бдительности птицы, важным для потенциального хищника. Однако оно выдаётся автоматически, независимо от наличия хищника на горизонте. См. Randler C. Is tail wagging in white wagtails, Motacilla alba, an honest signal of vigilance?// Animal Behaviour. 2006. V.71. №5. Р.1089-1093.
[37] См. Гомелюк В.Е., Тупоногов В.Н. Географическая изменчивость полового поведения трёхиглой колюшки// Поведение животных в сообществах. Мат. III Всес. конф. по поведению животных. М.: Наука, 1983. С.260-261.
[38] Эта демонстрация многофункциональна, как барабанная дробь дятлов или песня у воробьиных. Для птиц своего пола, с которыми конкурируют за партнера, она «означает» угрозу и стремление доминировать, для потенциального партнера — готовность поухаживать, для «нейтральных» особей — сигнал, сплачивающий чаек в «клубах» и других группах. Долгий крик — самый сложно структурированный полимодальный сигнал (вокальный и визуальный одновременно) чайковых птиц (сем.Laridae), в менее ритуализированной форме присутствует и у более примитивных поморников (сем. Stercoraridae). Включает определённую последовательность звуков, сопряжённых с серией глубоких поклонов и задираний головы (часто с отведением крыльев), которые чередуются между собой. Общая амплитуда «долгого крика», скорость исполнения демонстрации и фиксированность выразительных движений данного ритуала видоспецифичны, из-за чего сходство-различия его облика значимы при установлении родственных связей чаек Larus spp. (Moynihan, 1958, 1962). По многоэтапности и сложности координации последовательностей визуальных и вокальных сигналов животного, реализуемых параллельно друг другу, долгий крик сопоставим с «триумфальной церемонией» у гусей, обсуждаемой К. Лоренцем в «Агрессии».
[39] Как в целом, так и на каждом этапе последовательной реализации «крика».
[40] Они создают разнотипные «помехи» коммуникации, в общем случае не только не «смазывающие» специфические формы сигналов, но даже, наоборот, «подчёркивающие» их, см. ниже.
Автор — Владимир Фридман, для «XX2 ВЕКа».
Читайте предыдущие части серии:
20 фактов об инстинктах. Часть 1