Введение
Добрый день. Меня зовут Лучин Юрий Николаевич. Я хотел бы сегодня представить Вам свои мысли по работе головного мозга человека под общим названием “нейрофизиология и здравый смысл”. Сначала несколько слов о современной нейрофизиологии. По-моему мнению современная нейрофизиология слишком сильно упирает на различного рода лабораторные опыты, исследования и достаточно прикладные моменты и совсем немного времени уделяет общему осмыслению ситуации, общему пониманию принципов работы головного мозга. В своём выступлении я постараюсь восполнить этот небольшой пробел в современной нейрофизиологии. Это наука достаточно сильно переплетается с науками, которые находятся на граничных с ней участках, а это: генетика, эволюционистика, психофизиология. Без знания этих наук невозможно точно и в полном объёме разобраться в работе головного мозга. Но эти науки, перечисленные мной, также достаточно далеки от реальности, от реального понимания вещей. И поэтому перед тем как разобраться в работе головного мозга мне пришлось достаточно большое время потратить на то, чтобы разобраться и в генетике, и в эволюционистике, и в психофизиологии. И только благодаря тому, что мне удалось в них более-менее разобраться я смог после этого приступить к изучению работы головного мозга. Что нам даёт знание генетики? Знание генетики или того, что лежит в основе генетики, а это ДНК, даёт нам базу для понимания всего живого, что есть в этом мире, то есть, не зная базы, невозможно переходить к дальнейшему изучению других наук связанных с жизнью. Что даёт знание эволюционистики? Знание эволюции даёт нам понимание дороги, по которой идёт эволюция, как она идёт, какими темпами. Знание эволюции даёт нам понимание каким образом идёт развитие построения головного мозга. А что даёт знание психофизиологии? Оно даёт нам то к чему приводит работа головного мозга, то есть к поведению человека.
Методология
Изучение любой науки, в том числе и нейрофизиологии, всегда должно базироваться на трёх принципах. Первый принцип это здравый смысл. Второй принцип это понимание пути эволюционного развития мозга. Третий принцип это полное объяснение всех лабораторных опытов. Первый принцип здравый смысл. Это то, без чего невозможно изучение не только нейрофизиологии, но и любой другой науки. Объяснение любого компонента головного мозга, как и головного мозга в целом, всегда должно происходить с точки зрения здравого смысла. Приведу такой пример. Есть такое предприятие АвтоВАЗ и вокруг АвтоВАЗа, как подсказывает нам здравый смысл, будут располагаться только предприятия связанные с ним и в той или другой степени, задействованные в процессе этого производства. Вероятность того, что там будет обнаружена, допустим, академия художеств полностью отсутствует. Я привел этот пример для того, чтобы объяснить, что такое центральное серое вещество, которое располагается вокруг центрального спинномозгового канала. Современное объяснение немножко противоречит здравому смыслу. То скопление нейронов, которое находится вокруг водопроводного канала – это те нейроны, которые исключительно группируются вокруг канала для того, чтобы связать нервную систему с той жидкостью, которая находится в этом канале, то есть или для получения различных веществ из этого канала или для передачи их в этот канал. Всё что происходит потом с нейронами, которые сгруппированы вокруг центральной спинномозговой жидкости – это всего лишь следствие того, что нейроны находятся там и взаимодействуют друг с другом. Следующий принцип это понимание пути эволюционного развития мозга. Начиная с начала построения и по момент окончательного формирования. Знание этого пути обязательно для того, чтобы точно понимать значение тех или других ядер внутри головного мозга, тех или других элементов мозга. И третий принцип это полное объяснение всех результатов лабораторных опытов. Не так как обычно бывает, когда объясняется один показатель или одно следствие или один результат, остальные, допустим, пять игнорируются, те результаты, которые, к примеру, противоречат первому объяснению. И только полное объяснение всех результатов может претендовать на то, чтобы считаться объяснением действия того или иного нейрона, или ядра.
Параметры нервной системы
Потенциал действия в миелиновом волокне
Для того чтобы приступить к рассмотрению работы головного мозга нам надо будет ввести некоторые дополнительные параметры, потому что без этого невозможно понять, как работает головной мозг. Невозможно научиться читать, если человек не знает часть алфавита. Поэтому я ввиду несколько дополнительных понятий, первым из которых будет рассмотрен механизм работы движения возбуждения по нервной клетке, то есть потенциал действия. Для того чтобы понять, как осуществляется потенциал действия достаточно посмотреть на то, как проходит возбуждение по нервному волокну в клетках покрытых миелиновой оболочкой. Всё время движения возбуждения по нервному волокну отводится на движение ионов по ионным каналам, а между перехватами Ранвье всё осуществляется мгновенно, но мгновенно, в принципе, это невозможно, и здравый смысл подсказывает, что то, что мы считаем мгновенно – это есть просто движение электромагнитных волн со скоростью триста тысяч километров в секунду. И это по-новому позволяет нам взглянуть на проведение возбуждения в нервных клетках. Ведь современное представление о том, что движение возбуждения идёт электротонически, то есть через движение ионов, имеющих большую массу по вязкой субстанции внутри аксона или дендрита, оно не выдерживает никакой критики. И поэтому тот механизм, который я сказал, что проходит с помощью электромагнитных волн, но при этом современная наука как-то на этом не акцентируется, но здравый смысл нам подсказывает, что именно так это всё и происходит. Если в миелиновых волокнах всё происходит с помощью быстрых перескоков между перехватами Ранвье, то естественно, что и в безмиелиновых волокнах происходит точно такое же движение возбуждения, то есть движение возбуждения через распространение электромагнитных волн. Как это происходит? При открытом канале происходит движение или ток ионов через канал. Этот электроток создаёт магнитное поле, которое распространяется во все стороны. Но мы знаем, что магнитное поле действует только на движущиеся заряды, а не на статические и в окрестности там везде, естественно, будут статические заряды, но по-другому если рассматривать, что для того чтобы действие на статические заряды электрические магнитного поля надо чтобы это магнитное поле менялось и, действительно, рассматривая расположение ионных каналов, в которых находятся много во все стороны по оболочке, то есть мы приходим к выводу, что в каждом канале возникает маленькое магнитное поле. И так как магнитное поле возникающее, оно всё время меняется, то вот это изменяющее магнитное поле и заставляет двигаться статические заряды, которые находятся вне оболочки при распространении потенциала действия. И получается у нас система, когда движущие заряды создают магнитное поле, изменяющиеся магнитные поля начинают двигать следующие статические заряды, открывать ворота ионные каналы и дальше двигать поле. И такое распространение потенциала действия это самое простое, что можно изложить, видя вот эти все параметры.
Магнитное поле в нейроне
Ещё раз повторю, что магнитное поле возникает только при движении заряженных частиц и действует только на движущиеся заряды, на статические заряды может действовать и заставить их двигаться только изменяющее магнитное поле. Отсюда следует, что ионные каналы более-менее равномерно распределены по мембране. Возбуждение движется по волокну не только с помощью потенциала действия, потому что каждый отдельный ионный канал во время движения заряженных частиц создает вокруг магнитные поля, которые, изменяясь, заставляют двигаться заряженные частицы на расстоянии своего воздействия. Это означает, что у нейрона постоянно будут двигаться (бегать) волны. Дальше. Магнитное поле направленно всегда перпендикулярно к траектории движения заряженной частицы. Это означает, что направление развития магнитных сил будет идти влево, вправо по волокну. Направление вектора магнитной индукции ионного канала жестко закреплённого на мембране будет только вниз или вверх, для натрий каналов оно будет вниз. Вектор магнитной индукции определяет силы, действующие на токи или движущие заряды в магнитном поле. Магнитное поле подчиняется принципу суперпозиции. Если магнитное поле создаётся несколькими проводниками с током, то индукция результирующего поля есть векторная сумма индукции полей, создаваемых в каждом проводником в отдельности, а так как мы знаем, что плотность потенциал зависимых каналов в дендритной мембране менее чем в 10 каналов на микрометр квадратный, тогда как перехват Ранвье в аксоне содержит до тысячи, двух тысяч быстрых натриевых каналов. Это означает, что при сложении магнитного поля будет результирующая такова, что увеличивающееся магнитное поле будет доставать до более отдалённых ионных каналов миелиновых нейронов до пяти перехватов Ранвье.
Волна гиперполяризации
Начало развития потенциала действия влияет на электроуправляемые калиевые каналы, при этом их более медленное открытие начинается через долю миллисекунды после открытия натриевых каналов. Выход калия запускается электрическим полем, которое увеличивается после деполяризации мембраны во время потенциала действия. И только потенциала действия, потому что работа одиночного натриевого канала не сильно изменит поляризацию. Но после активации калий каналов возникшее магнитное поле с направлением индукции из клетки будет, в свою очередь, воздействовать на соседние калий каналы, возникнет вторая волна – волна гиперполяризации. Если натрий каналы запускают волну деполяризации, то калий каналы запускают волну гиперполяризации. Волна гиперполяризации запускается также калий каналами запускаемыми медиатором ГАМК – гамма-аминомасляная кислота. Разница между временем открытия натрий каналов и калий каналов объясняется временем вхождения натрия в клетку.
Следовые потенциалы
Анализ работы магнитного поля позволяет нам раскрыть все белые пятна, связанные с работой нервной клетки с прохождением потенциала действия. Самое большое белое пятно это следовые потенциалы. Рассматривая действие ионных каналов, невозможно представить, что с начала ионы, допустим, натрия двигаются в клетку, потом отдыхают и потом снова начинают двигаться. Здравый смысл говорит о том, что второе начало движения ионов натрия оно обусловлено посторонним воздействием и как ранее было сказано, что это воздействие является воздействием магнитного поля. В безмиелиновых волокнах всё просто: сначала наступают отрицательные следовые потенциалы, потом положительные следовые потенциалы, потому что натрий каналов больше, чем калий каналов. С ионных каналов на большем расстоянии приходит воздействие образованного магнитного поля, больше магнитная индукция. В миелиновых нервных волокнах следовые изменения потенциала имеют сложный характер. Следовый отрицательный потенциал часто сменяется следовым положительным потенциалом, затем иногда развивается новая электроотрицательность и лишь после этого происходит полное восстановление потенциала покоя. Рассмотрим следующую таблицу.
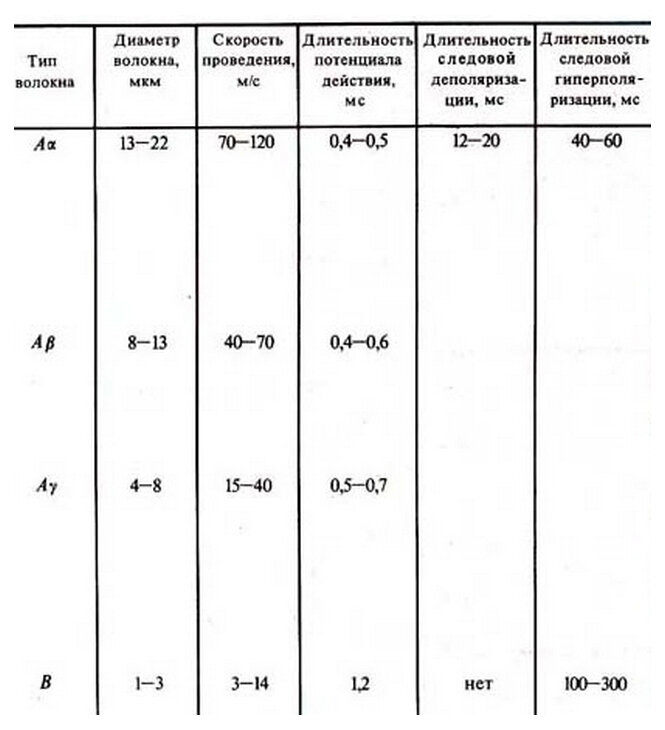
Здесь указана длительность следовых потенциалов для миелиновых волокон. Тип волокна А альфа отрицательные следовые потенциалы имеют продолжительность 12-20 миллисекунд, положительные следовые потенциалы для этих волокон имеют продолжительность 40-60 миллисекунд. Для волокон А бета, А гамма, А дельта отрицательные и положительные следовые потенциалы отсутствуют. В волокнах В отрицательные следовые потенциалы отсутствуют, а положительные имеют продолжительность от 100 до 300 миллисекунд. Такое развитие событий объясняется достаточно просто. В ближайших перехватах Ранвье просто не доходит магнитное поле, тогда следовые потенциалы отсутствуют, а если следовые потенциалы присутствуют, это означает, что с ближайшего перехвата Ранвье, там где прошёл потенциал действия доходит магнитное поле и, соответственно, вызывает или отрицательные или положительные следовые потенциалы. Если в миелиновых волокнах возникает чередование отрицательного, потом положительного, потом снова отрицательного следового потенциала – это означает, что второй следовый потенциал, магнитное поле доходит со второго перехвата Ранвье.
Электрическое поле в нейроне
Рассмотрим структуру потенциала действия. Сейчас трактовка такова: под влиянием короткого деполяризирующего стимула открывается часть потенциал зависимых натриевых каналов, что делает мембрану более проницаемой, ещё дальше смещает мембранный потенциал по направлению к равновесному натриевому потенциалу. В результате такой положительной и обратной связи открывается ещё больше натриевых каналов и так продолжается до тех пор, пока не возникнет потенциал действия. Один вариант и второй вариант, что изначально действуют медленные натриевые каналы, а потом с определённым временем подключаются быстрые натриевые каналы. Это очевидно совсем не так, потому что если бы было так, то форма потенциала действия имела бы после медленной деполяризации ещё участок напоминающий гиперболу, потому что не все быстрые натриевые каналы одновременно откроются, будет какая-то задержка. А если посмотреть на форму, то там сразу идёт прямой участок – прямая линия это означает, что открыты сразу и все натриевые каналы. Такое возможно только тогда, когда есть один большой участник. Вот этим одним большим участником и является электрическое поле. Электрическое поле, которое сначала двигает мембрану потенциал вверх с момента начала потенциала действия, а потом, изменившиеся электрическое поле двигает мембранный потенциал вниз с помощью калиевых каналов. Дадим определение потенциала действия. Потенциал действия происходит тогда, когда к изменяющемуся магнитному полю подключается электрическое поле. Потенциал действия невозможен без участия электрического поля. В толстых волокнах, когда сразу наступает потенциал действия, сначала действует только магнитное поле, потом присоединяется и электрическое поле. На форме потенциал действия это присоединение не будет сказываться. Прямая линия потенциала действия это все ионные каналы открыты, через них проходит максимально возможное количество ионов натрия вверх и калия вниз. Если какие-либо каналы станут закрываться или будут не открыты, а потом открыты, то линия потенциала действия будет наклоняться в горизонтальную сторону. Прерывание распространения потенциала действия всегда будет следствием увеличения расстояния между одиночными натриевыми ионными каналами. Угол наклона прямой линии фазы деполяризации потенциала действия более вертикальный, чем фаза реполяризации свидетельствует, что количество натрий каналов и ток ионов натрия выше, чем калия.
Миелиновая оболочка
Миелиновая оболочка является следствием прохождения большого числа потенциалов действия по нервному волокну. Часть ионов кальция доходит до глиальной клетки. Входящий кальций внутри олигодендроцитов активирует работу агранулярного эндоплазматического ретикулума, который формирует мембрану клетки. Строящаяся мембрана будет увеличиваться и свободным концом будет наматываться на ближайшее нервное волокно. Чем больше будет выделяться кальция, тем больше будет слоёв миелина или, что то же самое, увеличение количества потенциалов действия, увеличение амплитуды потенциалов действия будет увеличивать число слоёв миелина. На определённом уровне активности нейрона число слоёв будет постоянным. Демиелинизация происходит тогда, когда резко снижается количество потенциалов действия и их амплитуда. Кроме ионов кальция на увеличение количества слоёв миелина влияет аденозин – это вещество, в которое превращается молекула АТФ после отщепления от неё остатков фосфорной кислоты, но никак единственный активатор, а как вещество, способствующее активации всех процессов происходящих после использования энергии аденозинтрифосфата.
Перехват Ранвье
Образование перехватов Ранвье объясняется следующим образом. Созданные ионные каналы транспортируются аксонным транспортом. Проходящий потенциал действия изменяет количество кальция внутри клетки. В результате воздействия кальция на аксонный транспорт происходит освобождение ионных каналов, которые диффундируют клеточной мембране. И с помощью конечной сигнальной молекулы входят в мембрану и закрепляются в ней. Ионные каналы не встраиваются в мембраны под миелином, потому что там нет всплесков кальция. Большое количество натриевых каналов в перехватах Ранвье является основой для создания большого магнитного поля и большей скорости проведения возбуждения. И поэтому эволюционное предназначение миелиновой оболочки не в создании изолирующего слоя мембраны, а в группировании ионных каналов в одном месте для создания, как можно большего магнитного поля и большой скорости проведения возбуждения, что привело в конченом итоге к формированию морфологически сложного организма.
Рост дендритов
Показано, что нейроны с более сложной морфологией большие и разветвленные дендритные дерева, больше веток и шипиков, характеризуются пачечной импульсацией, тогда как нейроны с более простой морфологией имеют тенденцию к тоническим спайкам. Это объясняется тем, что во время прохождения потенциала действия по нейрону будет происходить рост дендритов. Рост дендритов будет происходить в следующей последовательности: во время потенциала действия происходит выброс кальция, который активирует работу эндоплазматического ретикулума по построению мембраной оболочки из фосфолипидов. И поэтому эндоплазматический ретикулум найден в дендритах клеток, но не найден в аксонах. Эндоплазматический ретикулум будет находиться в местах ветвления дендритов. Диаметр ветвления каждого следующего дендрита меньше предыдущего, вследствие того, что эндоплазматический ретикулум находится внутри дендрита, его размер будет меньше. Диаметр последней ветви дендрита пирамидного нейрона значительно меньше, чем в родительской ветке. Это означает, что достигнуто наименьшее создаваемое эндоплазматическим ретикулумом мембранное кольцо. Более тонкие ветви дендритов протяжённее, чем более толстые и показывают, что единицей отсчёта здесь является не длина дендрита, а площадь поверхности. Ветви аксонов образуют тупые углы и имеют диаметры такие же, как и родительские, потому что создаются эндоплазматическим ретикулумом находящемся в теле клетке. Рост дендритов нейронов будет идти по прямой линии из равномерного образования липидов по окружности, а так как в мембране располагается небольшое количество интегральных белков, то дендриты будут очень гибкими и препятствие на пути распространения приведёт к изменению направления роста.
Возбуждение дендритов
Знание работы магнитного поля в нейроне позволяет расставить все акценты в проведении возбуждения по дендриту на допороговом уровне. Чем меньше плотность ионных каналов, тем меньше скорость одиночной волны. Очевидно, что в самых тонких волокнах она будет самой маленькой. При маленькой плотности ионных каналов одиночный спайк не дойдёт до тела, он перейдёт в небольшую допороговую волну. Обязательно для каждого нейрона при переходе с дендрита на тело будет уменьшение скорости проведения потенциала действия или его полное прекращение из-за того, что величина магнитной индукции будет уменьшаться вследствие увеличения расстояния между диагональными, относительно оси нейрона, ионными каналами, из-за увеличения размеров мембранной поверхности по ходу потенциала действия. Соответственно, до достижения потенциала действия максимального диаметра клетки высока вероятность его прекращения без помощи тормозных нейронов. После перехода потенциала действия через максимальный диаметр клетки самопроизвольно он не прервется.
Аксонный холмик
Раннее сказанное позволяет нам достаточно просто определит причину возникновения потенциала действия в нейроне. Понятно, что нет никакого аксонного холмика со специальной мембраной. Просто, когда волна возбуждения на допороговом уровне движется от максимального диаметра клетки к аксону, магнитная индукция все время будет увеличиваться, соответственно, будет происходить все большая деполяризация из-за уменьшения расстояния меду диагональными натриевыми ионными каналами по ходу прохождения возбуждения вдоль оси нервной клетки. И в какой-то момент увеличивающаяся деполяризация превысит критический уровень и развивающаяся электрическое поле сформирует потенциал действия, он может возникнуть в любом месте от самого начала аксона до места выше по телу клетки.
Форма нейронов
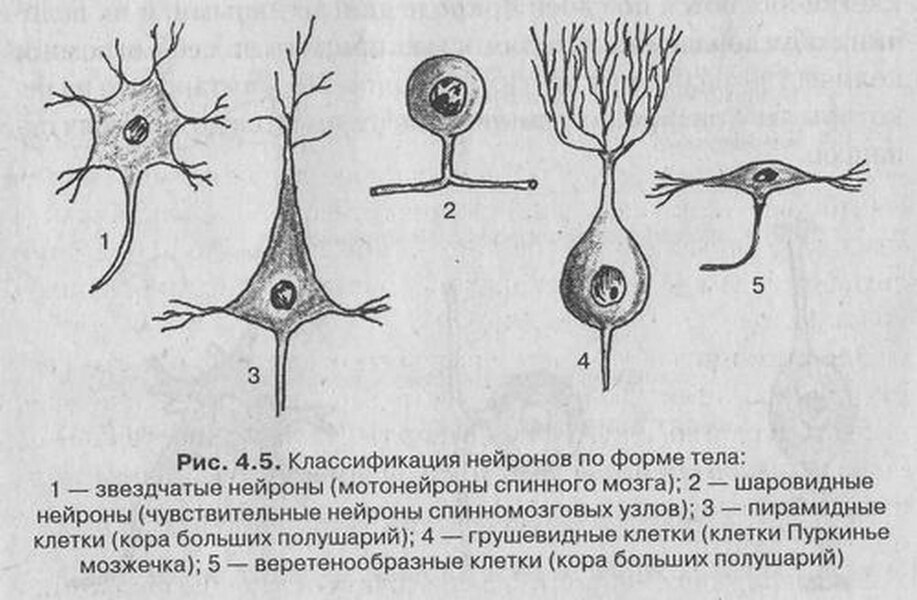
Форма тела не пирамидных нейронов варьируется в зависимости от типа, в основном имеет овальную форму. Рассмотрим каким образом меняется форма тела. Клеточная мембрана – это эластическая молекулярная структура, состоящая из белков и липидов, в основном фосфолипидов. Белки чаще пронизывают мембрану насквозь, например ионные каналы. При увеличении интенсивности работы нейрона, то есть проведения потенциалов действий происходит улучшение метаболизма, увеличение образования новых ионных каналов, и соответственно, встраивание их в мембрану, и чем больше ионных каналов будет встраиваться в мембрану там где больше проводится потенциалов действия, а там где в основном идут только волны возбуждения – там будет меньше ионных каналов. Так как клеточная мембрана двухслойная и она огибает клетку похожую на шар, то расстояние между верхними гидрофильными участками будет больше, чем между нижними и встраивание ионных каналов с помощью сигнальной молекулы приведёт к уменьшению этой разницы. Чем больше будет ионных каналов, тем больше будет выпрямляться окружная мембрана, то есть она будет стремиться к прямой поверхности. Отсюда легко заметить, что в луче звездчатого нейрона большая плотность ионных каналов, чем в самом теле и чем по мере перехода луча в тело будет наблюдаться уменьшение плотности ионных каналов. В пирамидном нейроне конусовидная форма со стороны апикального дендрита показывает, что здесь идёт большой поток возбуждения на допороговом и послепороговом уровне. Треугольная форма пирамидных нейронов показывает, что поток возбуждения со стороны апикального дендрита гораздо больше, чем со стороны аксона. Это может быть, например, когда аксон подтормаживается канделябровым нейроном. Если пирамидный нейрон имеет форму ромба, то это значит, что возбуждение вверху клетки сравнима с тем, которая снизу. Это когда каждый поток импульсации приводит к образованию потенциала в аксоне. Веретеновидные нейроны получаются в результате прохода неизменяемого потока возбуждения через нейрон, когда потенциал действия проходит через нейрон дальше по аксону или также проходит волна возбуждения на допороговом уровне. Знание этих принципов позволяет объяснить любую форму нейрона.
Апикальные дендриты
Наклон апикальных дендритов. Следствием большого количества ионных каналов в апикальном дендрите будет его очень большая жесткость на изгиб. Это видно в третьем слое новой коры, где апикальные дендриты пирамидных нейронов третьего слоя расположены под углом к вертикали, потому что они группируются в пучок. Тело пирамидного нейрона вслед за апикальным дендритом также наклоняется в ту же сторону.
Микроглия
Микроглиоциты расположены в коре равномерно и притом в шахматном порядке, наподобие протоплазматических астроцитов. Каждый микроглиоцит имеет свой район действия. Область распространения отростков одного микроглиоцита не переходит на территорию смежного микроглиоцита. Каждый микроглиоцит со своими отростками связан с определёнными нервными и глиальными клетками и сосудами, расположенными в области их разветвления и как бы объединяет все эти элементы. После специализации микроглиоцита или в нейрон, или в астроцит, или в олигодендроцит происходит размыкание уже не нужных ему связей. Микроглиоциты различных слоёв коры головного мозга отличаются друг от друга, так микроглиоцит, расположенный в первом слое сравнительно меньше микроглиоцитов, расположенных более в глубоких слоях, но по степени разветвления не уступает им, то есть величина микроглии в разных слоях коры регулируется генетически.
Дендро-дендритический синапс
Знание принципов эволюционного развития нервной системы позволяет объяснить такое явление, как дендро-дендритический синапс. Установлены между клетками синапс рабочий, то есть функционирует, если бы это было не так, то он просто бы исчез, как нервные волокна после рождения. Дендрит в отличие от аксона не обладает везикулами с нейромедиаторами. Сейчас понятно, что передаточным звеном является изменяющееся магнитное поле, причём передаваться может или волна деполяризации, или волна гиперполяризации. Основное условие – она должна быть достаточно велика, чтобы перейти через дендро-дендритический синапс, то есть работающий синапс не исчезнет.
Ритмы ЭЭГ
Рассмотрим ритмы ЭЭГ. Дельта-ритм имеет частоту 1-3 Гц и амплитуду 200-300 мкВ, тета-ритм имеет частоту 4-7 Гц и амплитуду 100-200 мкВ, альфа-ритм имеет частоту 8-12 Гц и амплитуду в диапазоне 45-70 мкВ, бета-ритм имеет частоту 16-30 Гц и амплитуду 10-30 мкВ, гамма-ритм имеет частоту 30-90 Гц и амплитуду колебаний 8-12 мкВ. Несложно заметить, что произведение частоты на амплитуду во всех ритмах составляет приблизительно одно и то же число, назовём это число базовой цифрой сознания. Причём у каждого человека оно будет не таким, как у остальных людей, его можно искусственно поднять с помощью активированных систем, например, ацетилхолиновых нейронов или опустить, но в среднем оно будет одним и тем же. Зависит оно от количества таламических регулярно разряжающихся, в основном, в неокортексе нервных клеток, как пирамидных, так и других. Причём возбуждаться будут изначально исключительно пирамидные нейроны через терминали апикальных дендритов в первом слое коры и дальше по волокнам и различным контактам. Между нейронами возбуждение будет расходиться по коре. Волны будут идти на допороговом уровне. Это импульсация не имеет ничего общего с таламической специфической информацией на звездчатые нейроны четвертого слоя неокортекса. Отрицательные и положительные полисы расшифровываются следующим образом. Отрицательный полис означает, что движение волн по нервным волокнам будет от точки отводящего электрода во все стороны и вглубь коры. Причем это будет максимум ухода волн из под электрода. Нейтральный уровень это когда количество ухода волн равно количеству прихода. Положительный полис, когда все волны идут по направлению к электроду, от нижних слоёв коры, в основном вверх. Почему именно с терминальных синапсов апикальных дендритов пирамидных нейронов начинаются волны возбуждения в коре? Знание работы генетических механизмов позволяет объяснить это. Терминальные синапсы апикальных дендритов в первом слое в 3-4 раза крупнее боковых и содержат по три митохондрии, в то время как боковые совсем не содержат митохондрий. Кроме того сам шипиковый аппарат в конечных разветвлениях апикальных дендритов значительно крупнее, чем на стволе дендрита и содержит по четыре митохондрии. Все остальные шипики содержат всего по одной митохондрии. Синаптические аппараты на теле клетки и на начальных участках дендрита, где нет шипиков – меньше терминальных синапсов и содержат по одной митохондрии. Эволюционно выработан механизм, что на внешние воздействия клетка отвечает выработкой требуемых для ответа элементов и размещение их в нужных местах. Нахождение четырех митохондрий в шипиковом аппарате в терминали показывает, что суммарное возбуждение через него минимум в 4, а максимум более чем в 10 раз больше, чем в других синапсах. И поэтому эти нейроны имеют форму пирамиды, потому что по апикальным дендритам проводится основной поток возбуждения в коре. Вернемся к волнам. Приведу несколько лабораторных экспериментов. Поблизости от раздражаемого участка отводится главным образом отрицательный потенциал, а в более-менее отдаленных участках, а также в противоположном полушарии отводятся положительные потенциалы. Отрицательный потенциал обычно появляется здесь после положительной фазы. Начальная негативность в ответе на свет выражена преимущественно в поверхностных в первом, втором слоях коры. Погружение отводящего электрода вглубь коры шагами по одной десятой миллиметра вело к некоторому снижению амплитуды начально негативной волны ответа. На глубине три десятых миллиметра от поверхности коры происходило замещение начально негативной волны на начально позитивную. Дальнейшее погружение в кору сначала вело к увеличению, затем к выраженному уменьшению амплитуды начальной ставшей позитивной волны ответа. При углублении отводящего электрода под первый и второй слой сколько-нибудь значительно отрицательные потенциалы перестают отводиться. Умеренное раздражение, приложенное к коре в двигательной зоне конечности, приводит к появлению отрицательного потенциала, но не сопровождается сокращением мышц конечностей. Отрицательные потенциалы глубоких слоёв отражаются на поверхности, как положительные потенциалы. Если сочетать электрические раздражения коры в какой-либо первичной зоне с раздражением соответствующего рецептора, например, слуховую зону и слуховой рецептор, то с усилением отрицательного потенциала происходит угнетение спонтанной электрической активности наряду с угнетением первичных ответов от слухового раздражения. Ну и наконец, после приложения к коре раствора гамма-аминомасляной кислоты – ГАМК – стимул, вызывавший отрицательный потенциал часто начинает вызывать положительный. После аппликации ГАМК к пункту коры находящемуся на расстоянии нескольких миллиметров от пункта раздражения отрицательный потенциал часто устраняется без того, чтобы на его место заступил положительный потенциал. Послойное отведение показало, что обращение знака после приложения ГАМК происходит в том случае, если стимул вызывал возбуждение не только элементов поверхностных слоёв, но и глубоких. Если возбуждались только поверхностные, то ГАМК устранял отрицательный потенциал без обращения. Гамма-аминомасляная кислота здесь полностью останавливает движение волн в коре. А теперь о ритмах. Альфа-ритм – основной ритм. Бета-ритм показывает, как часто основной ритм режется тормозными воздействиями. Гамма-ритм – основополагающий ритм сознания. Дельта-ритм – это ритм минимума энергии и патологии. Тета-ритм – основной ритм восприятия и записи новой информации, когда все первичные проекционные положительные потенциалы складываются в единую сообщность, которая на графике электроэнцефалографии будет выглядеть, как нисходящая положительная прямая линия, например аккомодация глаза в зрительной чувствительности соответствует одному из первичных положительных потенциалов, когда все элементы зрительного механизма активированы, то формируется осознанное зрительное изображение. Более подробно вернемся к ритмам ЭЭГ при рассмотрении сознания. Надо понимать, что волны на ЭЭГ это подпороговые последствия распространения потенциала действия или пачки потенциалов действия, которые являются зачинателями этих волн, когда изначальный потенциал действия не может везде во всех своих разветвлениях нейрона инициировать потенциал действия, а только подпороговые волны. Зачинателем ЭЭГ волны всегда будет потенциал действия. В основе ЭЭГ волн будет изменяющееся магнитное поле. Действия электрического поля от потенциала действия будет малозаметно в самом начале волны. Поэтому ЭЭГ волны можно называть магнитоэхо или сокращённо магэхо. То о чём я до этого говорю сейчас трактуется, как синхронизация. Ну и соответственно, не сложно понять, что за явление, которое обозначается, как десинхронизация – просто волны возбуждения режутся с большой частотой или напрямую, или возбуждаются сети тормозных интернейронов. Основным тормозителем с определённой частотой является голубое пятно норадренергических нейронов.
Принцип Дейла
Нервная система устроена и функционирует немножко сложнее, чем это представляется современной науке, например принцип Дейла. Один нейрон, один медиатор может быть воплощён в конкретном нейроне, так и не может быть воплощён. Все зависит от требуемости в системной реакции большого количества нейронов, наличие в конкретном нейроне определённого набора требуемых веществ. Это происходит во время эмбриогенеза путем включения, выключения определённых наборов генов. Здесь надо вести речь о том, что должен быть один быстродействующий, другой продолжительно действующий эффект. Например, ГАМК эндорфин вместе на одном нейроне означает быстрое торможение, длительное подтормаживание. Другой вариант: глутамат энкефалин означает быстрое разовое возбуждение и потом торможение всей цепи. И определяться это будет, как уже было сказано, требуемостью скоординированной работы всей системы.
Мозжечок
В качестве введения к анализу рассмотрения работы головного мозга я хотел бы рассмотреть мозжечок сам по себе. Функции мозжечка я рассмотрю попозже, когда будем рассматривать движение, а сейчас я просто хотел заострить ваше внимание на том, для чего предназначен мозжечок. Для чего предназначен мозжечок видно по внешнему строению мозжечка. Сам по себе он практически, как внешняя копия полушарий головного мозга, но только гораздо меньше. Все объяснения, которые есть в современной науке, они все достаточно частные и для того, чтобы понять, все-таки, зачем нужен мозжечок организму надо просто привлечь к ответу здравый смысл. Любые изменения, взаимодействия работы нейронов в каких-либо системах в коре или в подкорке – они связаны с увеличением размером этой коры, увеличением размеров ядер в подкорке, но для ответа в эволюции никогда не привлекается подобие головного мозга, подобие полушарий. Одно подобие полушарий это и есть только в виде мозжечка. Здравый смысл даёт нам только один вариант ответа, что мозжечок сам по себе он не усложняет или не упрощает работу головного мозга, он просто дублирует её. Для чего нужно дублирование работы головного мозга, зачем? Обращаясь, опять же к здравому смыслу, можно ответить, что сама по себе работа головного мозга, она как бы работа в идеальных условиях, допустим, в невесомости, но мы находимся на нашей планете с определённой силой тяжести и здравый смысл нам подсказывает, что всё дублирование нужно только для того, чтобы компенсировать силу тяжести на нашей планете в работе головного мозга. Вот для этого и нужно всё повторение тех или других позиций в работе полушарий. Для этого и нужен мозжечок.
Сознание. Основные постулаты
Рассказав о нескольких простых понятиях, не о всех остальные будут выводиться по мере необходимости. Рассмотрим сейчас самое сложное явление, которое есть в человеческом мозге это сознание. Понятно, что неокортекс связан с корой. Понятно, что всё возбуждение или почти всё приходит в кору от таламуса. Известно, что самым многочисленным нейромедиатором в коре является гамма-аминомасляная кислота, а это тормозной медиатор. Приблизительная схема вырисовывается следующей. Многочисленная возбуждающая афферентация тормозится с помощью гамма-аминомасляной кислоты, а остаток выводится в качестве осознанных процессов. Известно также, что с момента стимула осознание приходит позднее в среднем на две десятые секунды, то есть требуется время для подготовки процесса осознания. Единственная схема, которая удовлетворяет всем этим особенностям состоит в следующем. Первичная афферентация приходит в четвёртый слой коры на паукообразные звездчатые нейроны с короткими аксонами с густыми разветвлениями аксона в области своих же дендритов. Такой эволюционный механизм сформировался для гарантирования последующей передачи возбуждения без предварительной активации на близлежащие пирамидные нейроны. Дальше пирамидные клетки передают возбуждение из неокортекса в различные участки мозга для включения системы сознания для восприятия данного первоначального стимула. Получателями здесь являются специфический таламус, неспецифический таламус, мамиллярные тела, субикулум и другие. В результате комбинированная информация приходит сюда же в четвёртый слой коры на пирамидный нейрон, с которого и начинается все процессы осознания стимула. Причём этот пирамидный нейрон и первичная звездчатая паукообразная клетка имеют совершенно идентичную модальность. Для того, чтобы быстро среагировать этот пирамидный нейрон и получает постоянное предварительное возбуждение с частотой альфа-ритма на энцефалограмме. И именно для этого в синапсах терминалях апикальных дендритов находится много митохондрий, что без знания генетики, конечно, невозможно объяснить. Переводя на бытовой язык это похоже на чайник, стоящий на слабом огне, с температурой воды порядка 90 градусов. Так разными способами эволюцией решаются вопросы гарантированной передачи импульсации. В звездчатых клетках – многочисленными аксодендритными синапсами, в пирамидных нейронах – за счёт предварительного возбуждения через апикальные дендриты. Звездчатая клетка здесь выступает в качестве этакого рубильника. Кроме всего прочего отсюда вытекает ещё один вывод. Модальность сигнала это качество конкретного нейрона и обусловлено конкретной комбинацией генов в ДНК, а не частотой и амплитудой потенциала действия, как до сих пор пытаются всё это объяснять. Частота и амплитуда увеличивают скорость доставки или силу воздействия, а не изменяют модальность сигнала. И разница между пирамидным нейроном и звездчатым одной модальности в том, что к одному подходит периодическая импульсация на частоте альфа-ритма, а к другому не подходит.
Неокортекс
Рассмотрим некоторые особенности построения неокортекса. Новая кора состоит из вертикальных миниколонок, центром которых или основой является отличающееся от остальных модально-территориальные сигналы, приходящие из таламуса. Вокруг проекционных простых зон находятся более сложные поля, например, в зрительной коре вокруг поля 17 находятся поля 18 и 19. В миниколонках последних центрами являются сформированные сигналы из ограды. И поэтому легко заметить, что ограда это орган подготовки к осознанному восприятию в окружающем нас мире разных видов чувствительности. Четвёртый слой – это слой входа в кору, пятый слой – выход из коры. Сформулируем принцип построения неокортекса. Изначально образуется базовый неокортекс с последующим нарастанием слоёв и подслоёв – вверх и вниз. Базовый неокортекс – это рядом расположенные миниколонки, центрами которых являются специфические модально-территориальные сигналы, которые приходят к звездчатому нейрону или уходят из коры от пирамидного нейрона. Обязательным условием здесь является то, что источники прихода сигнала и получатели входящего сигнала находятся вне неокортекса. Миниколонка по вертикали может занимать один слой – приёма или передачи сигнала, а может занимать два слоя – приёма и передачи, но обязательным в этом случае будет полная идентичность сигнала. Базовый принимающий слой неокртекса, а это 4-й слой формируется как за счёт всей чувствительности афферентной, то есть все рецепторы различных тканей и органов представлены в нём, так и за счёт нейронов ограды. В расположении чувствительных афферентов различных органов в 4-м слое неокортекса, которые сейчас еще не найдены, несложно определить. Они будут на прямой линии, которая проводится от органа через таламус и на противополжно полушарную кору. Выход с пятого слоя неокортекса будет на двигательные и вегетативные образования. Причём большие блоки сознательных выходов двигательных и вегетативных располагаются, соответственно, в полях 4, 6 и 11, 12. Кроме них будут небольшие вкрапления двигательных и вегетативных эфферентов за центральной бороздой, о причине и расположении которых будет рассказано далее.
Слои неокортекса
Если рассматривать любую кору в мозге, то видно, что сверху кора всегда располагается слой толстых и тонких волокон, связывающих нейроны коры с подкорковыми образованиями. Изначально волокна идут вокруг работающих нейронов на границе с другими образованиями. Несложно по аналогии отметить, что и наружная полоска Белларже и внутренняя полоска Белларже, и полоска Бехтерева – это все скопления волокон бывших в своё время конечными вокруг работающих нейронов базового неокортекса. Направления разрастания коры будет вверх и вниз от границы между 4-м и 5-м слоем коры. Наружная полоска Белларже – это наружные волокна базового принимающего слоя, а внутренняя полоска Белларже – это наружные волокна базового выходящего слоя. Полоска Бехтерева – это наружные волокна эволюционно более позднего третьего слоя коры. Третий слой коры – это основной ассоциативный слой неокортекса. Например, звуки, кодируемые приходящими в четвёртый слой височной коры, в третьем слое группируются в слова. И наконец, чтобы понять функции второго слоя коры, достаточно вспомнить, что во второй слой посылают аксоны, нейроны, подушки, и что у китов, подводных млекопитающих, нет второго слоя неокортекса, как у наземных млекопитающих, то есть у них очень слабое зрение. Отсюда выходит, что второй слой коры это окружающий нас мир в нашем зрительном поле, а нынешний первый слой обслуживает, по большей мере, этот второй слой.
Борозды и извилины
Рассмотрим исчерченность коры бороздами и извилинами. Центральная борозда – это линия, разделяющая весь неокортекс на два принципиально разных функциональных блока – спереди блока управления и сзади блока предмета управления или субъекта и объекта. Блок управления развивается на основе двигательных и вегетативных выходов, это сейчас сформировано, как поля 4, 6 и 11, 12. Блок предмета управления это вся афферентация оценивающая, как внутренние показатели, так и внешнюю среду в окружающем нас мире. Спереди борозды и извилины чаще направлены вдоль продольной оси мозга, а сзади поперечной. Расположение нейронов вдоль прямой борозды говорит о том, что они выполняют похожие функции, но различаются территориально. А еще линейное расположение нейронов говорит о одновременности появления их в неокортексе и в эволюции. Соответственно, круговая извилина, выполняющая похожие функции появилась одновременно после образования центра этого круга в коре мозга.
Лобная кора
Так как неокортекс это часть сознания, то понятно, что поле 4 коры – это осознанное возбуждение конкретных двигательных клеток. Премоторная кора в поле 6 – это коррекция осознанных движений групп мышц через зубчатое ядро мозжечка, оптимизирующее влияние силы тяжести через предварительную импульсацию за несколько миллисекунд до начала движения. Дополнительная моторная кора в поле 6 состоит из двух частей – восприятия и совместной работы с неостриатумом, они будут рассмотрены позже в соответствующих разделах. Полем 8 кодируются механизмы для осознанного восприятия различных видов чувствительности, например глазодвигательных механизмов, у собак это и механизмы управления поворотом ушей на звук и так далее. Поле 9 – это комбинированная долговременная память организма. Это изображение квартиры, улицы, конструкция автомобиля, стихотворения, кинофильмы. В поле 9 собраны разнообразные последовательности активации групп нейронов или только нейронов, разбросанных по всему неокортексу. В поле 9 также как у человека сильно развито у человекообразных обезьян. Рядом находящееся поле 10 у этих обезьян почти не развито, а у человека оно такое же большое. Этим полем 10, прежде всего, и отличается человек от обезьяны. Отвечает оно совместно с хвостатым ядром за так называемую кратковременную память, правильней это следует назвать как подсознание. Начинает функционировать механизм подсознание сразу же. Сознание с подсознанием находятся в реципрокных отношениях. Когда человек идёт по улице и разговаривает, то разговор контролируется сознанием, а ходьба подсознанием – полем 10 коры и хвостатым ядром. Сам механизм подсознания будет рассмотрен далее. Изъятие информации из поля 10 осуществляется через рядом расположенное поле 46. Ёмкость кратковременной памяти у человека будет пропорциональна количеству нейронов в поле 10, а также увеличению активации с помощью таламуса, холинергической и дофаминергической систем. Около поля 46 располагается поле 47, которое выполняет похожие функции по отношению к долговременной осознанной памяти, находящейся в поле 9. Так как были уже затронуты основные управляющие поля в неокортексе, то осталось сказать, как осуществляется управление ими. Основной осознанный вход в неокортекс происходит через переднюю поясную кору, он отличается от неосознанного входа через зрение или слух тем, что зрительный образ не зависит от сознания, которое через определённое время просто осознается, а через переднюю поясную кору происходит процесс доставания из памяти и инициации различных осознанных процессов. Ну и осознанный выход из неокортекса осуществляется через заднюю поясную кору. В речедвигательном поле 44 префронтальной и латеральной коры речедвигательность является его вторичной функцией, а первично то, что в нём набору разнообразных звуков пристегивается смысл. Поле 45 комбинирует нейронами поля 44, формируя из отдельных слов фразы. Теперь после того как дан краткий анализ основным полям лобных ассоциативных зон, которые по заверениям современной нейрофизиологической науки и есть самое ценное, самое сложное и самое умное в человеке перейдём, действительно, к самым сложным, самым ценным и самым умным образованиям, то есть тем, которые появились до образования неокортекса, и которые были зачинателями появления новой коры, а неокортекс это инструмент, с помощью которого внешние стимулы преобразуются во внутренние осознанные.
Осознание. Зеркальные нейроны
Анализируя наступление осознания различных видов чувствительности, например: тактильные через приблизительно 140 миллисекунд, зрительные через 180 миллисекунд. И зная, что требуется определённое время для того, чтобы скоординировать действия различных элементов сознания, несложно сделать вывод о том, что единственным возможным вариантом сознания является порционное. Продолжительность одной порции сознания равна продолжительности одной фазы тета-ритма гиппокампа. Порция лимитируется временем необходимым для осознания стимула самого сложного вида чувствительности, самым длительным процессом осознания, а это зрение. Если бы это было не так, то мы бы не одинаково осознавали тактильные и зрительные стимулы, и время между их осознанием всё время увеличивалось бы. Здесь надо отметить основополагающий принцип сознания – принцип деления. Временной механизм сознания будет делиться до тех пор, пока из минимально возможных элементов не будет создаваться картина, полностью отображающая окружающую действительность. Согласно этого принципа сознание формируется из порций, но если бы этим и ограничивалось бы, то восприятие было слишком расплывчатым. Поэтому чтобы восприятие не было бы расплывчатым формируется новая структура ретикулярное ядро таламуса, которое делит сознание на еще более мелкие элементы на частоте около 40 Гц. Это более мелкий элемент сознания назовем ником, то есть ник это базовый кирпичик сознания. Он будет представлять этакую фотографию сознания. Информация для осознания поставляется двух типов – обонятельная и остальных видов чувствительности, приемники которых располагаются в неокортексе. Участки неокортекса, с которых поступает информация для осознания это вторичные соматосенсорные и моторные области коры, ретросплениальная кора и другие области. Характерной особенностью этих зон является более примитивная организация даже по сравнению с первичными полями сенсорных систем, а также более раннее эволюционное возникновение по сравнению, например, с ассоциативными зонами. В них находятся так называемые зеркальные нейроны. Импульсация этих нейронов означает, что в данный момент времени человек осознаёт то, что кодируют зеркальные нейроны. И когда человек смотрит на руку другого человека, это не означает, что он предполагает будущую программу действия этой рукой, он просто осознаёт то, на что он смотрит, то есть здесь наукой перепутаны причинно-следственные связи. Сначала проходят импульсы по нейронам кодирующим руку, а потом он осознаёт, что это рука. Легко заметить, что зеркального здесь ничего нет, просто эти нейроны должны обозначаться, как осознавающие нейроны. Также просто объясняется то, что эволюционно эти участки неокортекса более ранние. Зачем организму нужно осознание, если его невозможно осознать, и соответственно, нельзя им пользоваться. Разобравшись с зонами восприятия, и, проследив, куда направляют свои волокна осознавающие нейроны, находим места, где происходит процесс осознания – это гиппокамп, поясная извилина, миндалина, парагиппокамп, энторинальная кора. И так как таких мест осознания достаточно много, то можно сказать, что кроме того, что сознание порционное оно ещё и скачущее. И поэтому двусторонняя потеря гиппокампа, а это первый возникающий и центральный элемент осознавания, не ведёт к потере сознания человеком.
Гиппокамп
Рассмотрим строение гиппокампа. Гиппокамп состоит из большого количества одинаковых ламелл. Основой каждой ламеллы является модально-территориальный нейрон аналогичный центру миниколонки в неокортексе за центральной бороздой. Основной задачей ламелл является осознание сигнала приходящего в неокортекс и первичное комбинирование осознаваемых сигналов. Происходить это будет с помощью рекуррентных сетей и продольных связей в зоне CA 3 гипппокампа. Те ламеллы, которые будут активированы будут участвовать в процессе осознания. Пирамидные нейроны зоны CA 3 после активации функциональны до 12 секунд. Именно из этой зоны осуществляется оперативное управление осознанной деятельностью. Зубчатая фасция выполняет функцию выключения CA 3, то есть процесса осознания или поламилярно, или целиком всей продольной линии CA 3, в зависимости от поступающей информации, вида мышления, задействования других источников осознания. Информация после осознания по коллатералям Шаффера поступает в зону CA 1 для формирования комплексного восприятия с целью фиксации в память. Начало формирования комплексного восприятия будет происходить через нейроны начала образования блока информации, то есть в зоне CA 1 нет нейронов места есть нейроны начала. В зоне CA 1 несколько слоев пирамидных нейронов для поступления информации из CA 3, энторинальной коры, задней поясной извилины, соответственно. После осуществления процесса записи в память блока информации, последующее его извлечение из памяти на нейроны начала зоны CA 1 – бывшие нейроны места, будет сопровождаться пачечной импульсацией на тех слоях зоны CA 1, на которые пришла информация из разных мест осознания. Например, зрительная информация, вкладываемая в зону CA 1, будет служить зрительным фоном. Например, когда рассматривается какой-либо предмет в комнате, то в зоне CA 3 нейроны будут обозначать этот предмет, а в зоне CA 1 будет отображаться та часть комнаты, которая располагается по направлению оси взгляда. Причём выключенный свет не заставить прекратить импульсацию нейроны, кодирующие эту комнату в зоне CA 1. Хранится же изображение комнаты будет там же, где и текст романа “Идиот” – в поле 9 префронтальной коры. Следующая часть гиппокампа: субикулум предназначен для комплексного выключения, включения всех элементов мозга, требуемых для осознания данного стимула. Пресубикулум и парасубикулум осуществляют процесс межполушарного перехода сознания. Пресубикулум с их зерновидными нейронами осуществляет восприятие сознания, а парасубикулум с их пирамидными нейронами – передачу импульсации для инструментов сознания.
Энторинальная кора
Если до этого момента информация по разным структурам и полям была представлена очень кратко, потому что подробно об этом поговорим позже, то энторинальную кору рассмотрим сейчас подробнее, потому что она является самым изощренным образованием головного мозга, причём только некоторых млекопитающих самых морфологически сложных. Начнем с далека. В неокортексе в разных её частях находятся нейроны, активизация которых формирует сложные инграммы, например: мелодии, звуки слов, зрительные образы, лица и так далее. Понятно, что они образуются в результате восприятия человека, но сложно организованная нервная система мгновенно может провести только потенциал действия, но не образовать новый нейрон со сложным образом, причём только в конкретном месте и нигде больше. Поэтому эволюционно выработался механизм промежуточного хранения сложных образов в специально подготовленном месте. Это место находится в пятом слое энторинальной коры в виде плотно расположенных маленьких пирамидных нейронов. В эти пирамидки приходит комплексная информация по всем видам чувствительности, которая формируется в зоне CA 1 гиппокампа. Это похоже на видеоролик продолжительностью до 12 секунд со своим началом, а это первый потенциал действия приходящий в любой ламелле в зоне CA 1. Далее по очереди все потенциалы действия, поступающие в любую ламеллу CA 1, а оканчиваться он будет либо последним потенциалом действия в районе около плюс минус 12 секунд, либо окончанием тета-ритма в гиппокампе. Поэтому несложно определить, что тета-ритм – это ритм накапливания информации в блок памяти. Так называемый видеоролик может быть и значительно меньше 12 секунд. Для того, чтобы перезаписать этот сформированный блок информации на пирамидный нейрон в новую кору требуется повторить все манипуляции в гиппокампе. Но гиппокамп постоянно задействован в различных процессах и эволюционно начинает выстраиваться над пирамидками немного упрощенный вариант гиппокампа, когда продольно расположенная лента ретрокомиссулярного гиппокампа сворачивается в трубочку и в виде цилиндра надстраивается над пирамидками памяти, где 4-й слой крупных пирамидных нейроцитов выполняет функцию мгновенного стирания текущей информации по аналогии зубчатой фасции гиппокампа. Третий слой энторинальной коры – слой средних пирамид выполняет функцию CA 3 текущей оперативной осознанной деятельности. Второй слой аналогия CA 1 гиппокампа. Кстати, наличие ещё одного места оперативного осознанного управления третьего слоя энторинальной коры подталкивало некоторых любопытных нейрофизиологов изымать гиппокампы у людей и смотреть, что из этого получается, правда сейчас эта практика не пользуется большой популярностью. А в этих случаях будет заметно отсутствие возможности запоминания. Более тонкие нарушения не будут заметны без полного понимания работы гиппокампа. В медиальной энторинальной коре при заполнении пирамидок пятого слоя блоками информации возникнет потребность в новых пирамидках, она решается через дифференциацию микроглиоцитов и эпендимных клеток, маленькие пирамидные нейроны. При значительном превышении площади поперечного сечения пятого слоя над вертикальным цилиндром, новый цилиндр может появляться только в эмбриогенезе нового организма. Соответственно, эволюционно будет все время происходить увеличение количества вертикальных минигиппокампов в медиальной энторинальной коре. Так как минигиппокампов много в энторинальной коре, то задействование одного из них для осознавания будет происходить через блуждающее холинергическое пятно, связанная с нейронами направления головы в энторинальной коре. Если будет сильное холинергическое влияние, то осознаваемый видимый контур будет уменьшаться, и соответственно, он ничего не будет видеть совсем рядом. В конструкции глаза будет эволюционно происходить постоянное увеличение площади сетчатки глаза, когда каждому периферийному участку зрения или элементам сетчатки будет соответствовать свой минигиппокамп в медиальной энторинальной коре. Это происходит, потому что гиппокамп отправляет в зрительную память то, что находится по центру взгляда, что-то подобное нынешнему желтому пятну глаза человеком. Увеличение площади сферически расположенной сетчатки даёт панорамный эффект восприятия окружающего мира. Это даёт возможность осознавать большую площадь зрительного восприятия. Отсюда следует, что нейроны сейчас известные, как решетчатые являются вариантом нейронов начала, ранее известных как нейроны места. Этот путь рассуждений приводит к пониманию функции решетчатых нейронов довольно сложно. Более простой путь – это понять, почему решетчатые нейроны располагаются в виде гексагональной решетки. Просто только шесть окружностей одного диаметра можно разместить в округе ещё одной окружности этого же диаметра. А так как нейроны места очень похожи на нейроны решетки, то понятно, что наличие шести нейронов решетки вокруг седьмого означает наличие шести вертикально стоящих гиппокампов вокруг седьмого. Кстати, существует более десятка способов понимания функции решетчатых нейронов. Латеральная энторинальная кора гораздо меньше, потому что там хранится только обонятельная промежуточная информация. У энторинальной коры есть ещё одна функция, эта функция восприятия окружающего мира не связанного с собой, а наподобие видеофильма с определяющим зрительным участием. Ведь если есть участки оперативного осознанного управления, а это третий слой энторинальной коры, то неразумно было бы его не использовать. И поэтому вся зрительная и около зрительная информация начинает перекочевывать с гиппокампа на медиальную энторинальную кору, обладающую большими возможностями по сравнению с гиппокампом. По работе неокортекса известно, что воспринимаемая информация поступает на нейроны приёма, а это звездчатые нейроны, то поступаемая на эти нейроны второго слоя энторинальной коры направляется далее в третий слой для осознавания и дальше в зависимости от вида информации будет передаваться в другие участки своей коры: аммонова рога, субикулума и так далее.
Ядра таламуса. Септальная область
Ещё один участок, где находится оперативное осознанное управление это задняя поясная кора, где осознается вся информация касающееся самого человека, всего что связано с ним. Эта информация, как из энторинальной коры поставляется в зону CA 1 гиппокампа, для формирования комплексного субстрата для записи в память, но только в другой подслой CA 1. Процессы сознания активируются через передние таламические ядра: переднее медиальное, переднее вентральное, переднее дорсальное. Переднее медиальное ядро отвечает за инициацию осознанных процессов. Переднее вентральное – за конечное осознавание. Переднее дорсальное – за осознавание через зрительное поле. Передние ядра таламуса функциональны и очень похожи на механизмы действия после прихода специфической сенсорной информации на звездчатые нейроны четвёртого слоя новой коры в первичные поля. Они будут возбуждать пирамидные нейроны уже находящиеся под возбуждением со стороны других ядер таламуса. Соответственно, это будет давать гарантированную инициацию конкретных пирамидных нейронов. Ретикулярное ядро таламуса является основным в сознательной деятельности организма. Оно не тормозит специфические ядра таламуса, оно их возбуждение делит на части обычно 40 в секунду, так называемые ники. Эти срезы почти как в кинематографии поступают дальше для осознания в лимбическую кору. Эволюционно выработана такая высокая частота ников для функции сознания, чтобы не было возможности понять их статичность. Медиодорсальное ядро таламуса является основным узлом связи в текущей деятельности, как для образования временных соединений в поле 10 неокортекса кратковременной памяти или, что более правильно подсознания, идущих из хвостатого ядра, так и образования новых нейронов в лобных зонах и подкорковых ядрах головного мозга. Вся информация различных видов чувствительности связывается в медиодорсальном ядре по временному признаку и передается или для временного хранения, или для постоянного на конкретные нейроны, на так называемые сложные нейроны. Септум. Структура, которая инициирует тета-ритм в гиппокампе это септум. При тета-ритме происходит запись новой информации, запись осуществляется с определённых нейронов в зоне CA 1 гиппокампа, потом с этих же самых нейронов будет происходить воспроизведение зрительной информации в виде зрительного фона. Запись простых событий складируется на пирамидных нейронах энторинальной коры. Сложная объёмная информация переносится в поле 9 префронтальной коры. Мамиллярные тела являются координаторами связующим звеном всех структур для полноценной записи информации на новые клетки.
Обонятельный мозг
Теперь, когда стал понятен механизм сознания у высокоорганизованных животных, рассмотрим такую структуру, как обонятельный мозг. Сейчас есть разные точки зрения на это: атавизм, висцеральный мозг, регрессирующее образование, но почему-то внутри с одним из видов чувствительности – обонянием. В действительности, это большой механизм сознания, который может быть построен на самом развитом и сложном из видов чувствительности. У тех организмов, где он создавался это было обоняние. Сам по себе в чистом виде механизм сознание не сформировался бы, а потом после образования данного механизма им, немного видоизменив, были осознаны и остальные виды чувствительности, которые были размещены всем скопом за центральной бороздой головного мозга в виде неокортекса.
Эволюция. Основные постулаты
Тектум
Рассмотрим вторую составляющую трехопорной конструкции на чем держится современная нейрофизиология. Рассмотрим механизм появления самого сложного образования в этом мире – головного мозга, от момента возникновения нейрона. В основе развития нервной системы лежит генетически закрепленный механизм, когда нейроны разной направленности, соединяясь, осуществляют содружественную деятельность. Например, зрительный нейрон и нейрон, иннервирующий мышечную клетку, соединяясь, дают конкретный эффект, который закрепляется генетически, а соединение зрительного нейрона и обонятельного нейрона при соединении не даст никакого эффекта, и соответственно, эта связь через какое-то время будет разомкнута. Процесс повторения раз полученного соединения и записанного в ДНК будет повторен в эмбриогенезе. Появление нового вида рецептора или старого рецептора в новом органе приведёт к тому, что все они направляют свои волокна вверх, где будут устанавливать связи с подобными в конечном мозге. Взаимодействие в этом месте конечного мозга будет мгновенным, рефлекторным. Это высший интегративный центр морфологически несложных организмов, причём это будет единственный центр в конечном мозге, не может одновременно возникнуть где-то ещё похожий центр, называется такой центр тектум. Рассмотрим конструкцию тектума. Тектум состоит из большого количества слоёв. У амфибий в тектуме – 9 слоёв, у рептилий – 14 слоёв, у птиц – до 15-ти слоёв, а у млекопитающих только 7 слоёв. Вроде с увеличением морфологической сложности эволюционно шло увеличение количества слоёв, но вдруг у млекопитающих только 7 слоёв. Объяснение это достаточно простое, как и конструкция тектума. У млекопитающих развивается механизм сознание и часть тектума элиминируется, то есть та часть, которую компенсирует осознанная деятельность. Слоистое строение тектума, как уже было показано в коре сознания, то есть в неокортексе, опосредованно временными параметрами. Строительство тектума, как и неокортекса начинается с элементарных чувствительных нейронов и элементарных двигательных нейронов, расположенных рядом и от которых с течением эволюции вверх и вниз будут наслаиваться новые скопления нейронов. Каждый следующий слой будет являться следствием произошедших значимых эволюционных изменений в ряду организмов. Отличие от неокортекса будет только в том, что процесс образования неокортекса прошел достаточно быстро на уже готовых органах чувств с образованием достаточно большого количества приспособительных механизмов, а у тектума, который начал формироваться первым интегративным центром образование органа чувств, например глаза, шло эволюционное продолжительное время вместе с формированием тектума. И получается, что достаточно плоскостная зрительная структура неокортекса резко контрастирует с достаточно объёмными зрительными полями тектума. То что вверху тектума у млекопитающих находится различного рода зрительные механизмы означает, что они были эволюционно последними с различного рода взаимодействиями чувствительных органов. Соответственно, в самом внизу тектума будут механизмы возможных скоординированных действий мышечных групп. Равномерное чередование слоёв клеток тектума и слоёв волокон говорит исследователям о том, что эти волокна обслуживают, в первую очередь, именно нейроны, лежащие под волокнами в верхних слоях тектума и над волокнами в нижних слоях тектума. Объёмное расположение рецепторов чувствительных органов означает, что вверху будут эволюционно более поздние возникшие, а внизу – более ранние.
Ретикулярная формация
Главный вопрос, который нервная система должна постоянно решать в своём эволюционном развитии это как заставить импульсировать следующий нейрон, получающий возбуждение от предыдущего. И каждое следующее решение этого вопроса всегда приводило к эволюционному скачку в линии организмов. А первое и самое основополагающее это создание механизма позволяющее гарантированно возбудить следующий нейрон после рецепторного. Этим механизмом является ретикулярная формация. Ретикулярная формация образуется коллатералями чувствительных волокон идущих вверх в конечный мозг и коллатералями двигательных волокон идущих вниз. Образуется сеть, которая постоянно возбуждена, потому что организм живёт в постоянно меняющемся внешнем мире, с постоянным задействованием тех или других рецепторов и вынуждены рефлекторно на них отвечать. Соответственно, чем больше рецепторов, тем больше органов чувств; чем больше развит двигательный аппарат, тем гарантированее будет возбуждение следующего нейрона. Причём работе ретикулярной формации будет присуща характерная особенность – возбуждение на второй нейрон будет происходить из той зоны ретикулярной формации, куда будет первый модальный нейрон отдавать коллатераль. Таким образом второй модальный нейрон будет получать дополнительное возбуждение из того участка ретикулярной формации, которое гарантированно возбуждено, а не из невозбужденного участка.
Пейсмекеры
Следующим крупным, даже очень крупным шагом в развитии нервной системы, которая сделала возможным появление сложных морфологических организмов, явился возникший механизм самовозбуждения нейрона, когда соединились потенциал зависимый кальциевый канал и кальций зависимый калиевый канал. Это сделало возможным нейрону, и соответственно, нервной системе иметь гарантированное возбуждение и не зависеть от внешних раздражений. Организмы, функционирующие по принципу реагирования на что-либо дали жизни новым, которые стали сами инициаторами всего. Стали появляться органы чувств, построенные на постоянном возбуждении, организмы сами начали непрерывно двигаться. За счёт постоянного возбуждения у организмов начал функционировать метаболизм. Именно механизм самовозбуждения нейрона явился причиной появления на планете позвоночных. Этот механизм стимулирует активный оборот кальция, который вынимается из поступающей пищи, а потом излишки кальция связываются с ортофосфорной кислотой и получается фосфат кальция. Фосфат кальция образуется в местах активно работающей нервной системы. Именно активно работающая нервная система является инициатором возникновения костной ткани позвоночника, черепа и так далее. Раз появившейся механизм самовозбуждения через эмбриогенез следующих поколений был распространён на все существующие функции организма. Появились две структуры: в одном из которых таламусе на постоянное возбуждение были переведены все внешние воздействия, в другом гипоталамусе на постоянное возбуждение было переведено все возможные внутренние процессы. Причем таламус и гипоталамус не замещали какие-либо процессы, они добавляли новые.
Таламус
Изначально таламус создавался, как источник постоянного возбуждения направленного, как вверх в конечный мозг – это дорсальный таламус, так и вниз к мышечным клеткам – это вентральный таламус. Связи таламуса изначально создавались ипсилатеральными, то есть на постоянное возбуждение нейрона переводились пополушарно. Вентральный таламус позволил организмам, ранее передвигавшимся рефлекторно, по течению и так далее, приобрести способность к самоинициируемому безостановочному движению. Дорсальный таламус позволил резко активизировать и усложнить все процессы в конечном мозге у человека. Дорсальный таламус сейчас называется просто таламусом, а вентральный таламус, с навешенным на него большим количеством тормозных структур, называется субталамусом. Если бы субталамус постоянно не тормозился, то человек бы постоянно двигался, как акула. Основная функция таламуса возбудительная, и поэтому везде, где оканчиваются таламические нейроны будет возбуждение. Эволюционно на базе нейронов с периодическим самовозбуждением были выработаны несколько разновидностей. Проекционные таламические нейроны самые простые. Интраламинарные нейроны по сравнению с ними гораздо более сложные. Основная функция интраламинарных ядер – быстрый рост аксонов и дендритов, новых дифференцированных нейронов, за счёт подведения к нему максимально возможной импульсации, а это у интраламинарных нейронов выражается в частых пачках спайков до 80-ти Гц, внутри которых потенциалы действия следуют с частотой 800-1000 Гц. Соответственно, там где оканчиваются волокна интраламинарных нейронов – там происходит дифференциация клеток в нейроны, прежде всего микроглиоцитов, например, в ограде интраламинарные нейроны приходят в вентральную слуховую зону, а соответственно, в заднюю дорсальную зрительную зону они не приходят. С помощью ограды создаются новые видоизменённые модальные сигналы. Приход или не приход интраламинарных сигналов в ограду означает, что до сих пор у человека создаются акустические новые комбинации, а зрительные не создаются, потому что воспринимаемый зрением трёхмерный мир не меняется. Самые сложные таламические нейроны это нейроны медиодорсального ядра. Функция медиодорсального ядра таламуса – это временное или постоянное связывание всех возможных видов чувствительности на одном нейроне.
Гипоталамус
Гипоталамус, как высший центр вегетативной системы является наряду со зрительной системой, в первую очередь экстрапирамидной, местом приложения сил всех значимых нервных образований. Любой новый нервный субстрат, например новое ядро миндалины, обязательно будет оформлять свою деятельность в виде определённой двигательной активности и вегетативных реакциях. Причём сами эти направления жизнедеятельности будут обращаться друг к другу. Двигательная активность будет выражаться в вегетативных изменениях, а вегетативные изменения в различной двигательной активности. Функции гипоталамуса, как и таламуса будут рассмотрены далее. Скажем только, что сосцевидные тела никакого отношения к регулировке гомеостаза не имеют. Они выполняют функцию координатора механизма образования новых нейронов, точнее наделение их информационным содержанием, а находятся они в гипоталамусе, потому что появились они вместе с ядрами гипоталамуса в одно эволюционное время.
Ретикулярная формация. Торможение
Несложно предположить, что наряду с системой активации возбуждающих сигналов ретикулярной формации построена система активации тормозящих сигналов, которая по аналогии должна быть направлена вниз. Так как торможение сигналов в нервной системе также необходимо, как и самовозбуждение сигнала, потому что слишком сильное возбуждение может привести к разбалансировке. То естественно, что тормозящие процессы также должны быть гарантированы. Единственной системой гарантированной активации у простых организмов является ретикулярная формация. Поэтому в ретикулярной формации наряду с областями помогающими активировать поступающие возбуждающие сигналы появляются области помогающие активировать тормозящие сигналы. Запитываться они будут также как и с возбуждающими сигналами, за счёт коллатералей аксонов идущих вниз.
Модальность
Самый, наверное, веселый вопрос в современной нейрофизиологии это наделение той или иной модальностью импульсацию разных нейронов. Импульсация нейронов отличается по двум параметрам: частоте и амплитуде. Понятно, что невозможно всего двумя параметрами сформировать всё многообразие свойств любой модальности. Это как нельзя внутренний мир солдата оценивать по тому, как быстро он марширует или медленно, сильно он топает ногами или нет. Понятно, что только с помощью десятков разнообразных генов можно сформировать такие сложные ощущения, как вспышка света. Соответственно, все ощущения, такие как зрительные, акустические, тактильные, обонятельные и другие присущи только специализированным нейронам, в которых ощущения кодируются большими группами разнообразных генов.
Боль
Рассмотрим такую модальность, как боль. Для того, чтобы разобраться в таком понятии, как боль, прибегнем к помощи двух из трёх основ, на которых держится наука нейрофизиология это здравый смысл и знание механизмов формирования головного мозга. Известно, что зачатки системы ощущения боли есть даже у примитивных животных, они найдены не только у хордовых, но и у беспозвоночных, у которых нет сложного головного мозга, то есть искать ощущение боли надо на самом нижнем уровне организации нервной системы. У сложных организмов в головном мозге есть структуры, вовлеченные в это ощущение, но они больше как дополнение и для многообразия этого ощущения. Боль появилась одной из последних ощущений, которые есть у живых организмов, она не существуют сама по себе, как движение фотонов, колебания воздуха, наличие разнообразных молекул в воздухе, источников запаха. Для боли нужен определенный организм, покрытый оболочкой. Этому организму даже ненужно того большого разнообразия веществ, которые вызывают боль. И эволюционно у организмов появился механизм предупреждения об опасности, без чего невозможно дальнейшее морфологическое усложнение. Этим механизмом является сигнализация о разрыве внешней оболочки организма. У этого организма мог быть только один источник активации это ретикулярная формация и даже не сама ретикулярная формация, а принцип в её основе. Это принцип соединения большого количества волокон и наличия в нём определённого уровня возбуждения. Этакое колышащее море. Для этого нужно свободное нервное окончание, выходящее в покровы и наличие в этих окончаниях ионных каналов, активируемыми межклеточными воздействиями – сжатием или растяжением. И так как организм живой и покровы всё время изгибаются, то это обеспечивает постоянный поток возбуждения. Эти первично чувствительные нейроны посылают своё возбуждение в сеть, состоящих из большого количества специализированных нейронов боли, расположенных в первой пластинке маргинальных нейронов и частично в начале второй пластины, в начале желатинозной субстанции. Чтобы информация первичного чувствительного ноцицептора была гарантирована принята происходит разветвление аксона и он проходит вверх и вниз на один, два сегмента и даёт множество окончаний. Это и есть основная болевая импульсация. Располагаться болевые нейроны будут с самого края заднего рога, так как эта последняя модальность, находящееся в спинном мозге. И как было ранее сказано, формироваться она будет эволюционно последней, когда все другие модальности будут сформированы. Перед болевой зоной будут располагаться в части второй пластины и в третьей пластинке зона торможения болевой зоны, сформированная нейронами тепла, холода, прикосновения, давления и других приходящих волокон. Волокна, приходящие в эту зону идут вниз и вверх на три, семь сегментов, то есть в сумме до 14-и сегментов. Это нужно для гарантированного подавления в нормальном состоянии болевого возбуждения. Сюда приходит сверху волокна из ретикулярной формации, таламуса, центрального серого вещества для усиления активации подавления возбуждения. В конечном итоге получается, что вся система настроена на средний уровень болевого возбуждения со средним уровнем его подавления в норме выше, чем возбуждения. Любое отклонение будет приводить к увеличению боли, как низкий уровень серотонина, так и высокий приводит к появлению боли. В одном случае уменьшение присутствия тормозного медиатора серотонина – к увеличению возбуждения ноцицептора, в другом случае будет активация других болевых нейронов через выделение вещества П. Нейроны первой пластинки заднего рога, в которые приходит информация так называемого эпикритического пути, будут самыми крайними, так как они появились эволюционно последними и нужны они для осознанного восприятия болевого ощущения, то есть появились они с возникновением сознания. Эмоциональный компонент боли дают лобные зоны. Без лобных зон только в одной желатинозной субстанции боль будет чем-то похожа на скрежет, которая получается в виде генетически закрепленного видоизмененного аксонного транспорта, который резко усиливается при недостатке кальция и АТФ энергии в нейроны. Скрежет лучше ощущается, когда идёт от сети волокон, а не от одного. Система взаимодействия болевой сети и её торможения есть во всех основных болевых центрах: в спинном мозге, тройничном нерве, вентробазальном таламусе. Болевые ощущения, связанные с медиальным таламусом связаны с тем, что медиодорсальное ядро участвует в образовании новых связей, в том числе и болевых. Болевые ощущения, связанные с интраламинарными ядрами объясняются тем, что они обеспечивают рост аксонов и дендритов, новых нейронов, в том числе и болевых. Ну и последнее в этой теме. Отраженная боль при нарушениях работы внутренних органов, определяемая в спинном мозге объясняется тем, что эволюционно организму нужно было обозначить больные органы, и соответственно, эволюция не выдумывала новые системы обозначения боли, а направила волокна внутренних органов на нейроны уже готовой болевой системы. Понятно, что волокна от, например, сердца и желудка не будут конвергировать на одних и тех же нейронах спинного мозга, а на разных.
Ощущение удовольствия
Эволюционное развитие организмов на определённом этапе не было бы возможным, если бы наряду с обычным делением организмов с передачей генома не возник бы процесс комбинирования генома в будущих организмах. Это стало возможным, когда появились особи разных полов, и чтобы эти особи скрещивались должна была появиться причина, этой причиной стали сформировавшиеся нейроны, которые при прохождении потенциала действия обозначали бы существующему организму ощущения удовольствия. Это ощущение наступает, когда осуществляется системная реакция организма, и соответственно, взаимодействие большого количества нейронов, нацеленное на получение такого конечного результата, как удовольствие. Нейроны удовольствия или положительные нейроны, или плюс нейроны для краткости, составляют один из четырёх видов модально специфических нейронов, модальная специфичность которых закреплена генетически. И для воспроизведения модальности которых достаточно прохождение потенциала действия по нейрону. Остальные виды модальной специфичности нейронов работают только в комплексе со сложным прибором, например, зрение реагирует на изменение внешней среды, например акустические колебания воздуха, как основа звука. А наличие всего четырёх модально специфических простых видов нейронов вытекает из той подсказки современной науки, которую природа разместила на языке человека много лет назад, где горький вкус формируется на импульсации болевых нейронов, сладкий – на нейронах удовольствия, кислый и соленый – на тепловых и холодовых нейронах. Плюс нейроны находятся в прилегающем ядре. Они представляют из себя ацетилхолин содержащие интернейроны, очень крупные круглой или треугольной формы. Очень крупные нейроны, как мы уже знаем, это следствие больших потоков возбуждения, треугольная форма нейрона говорит о том, что к вершине треугольника подходит постоянная таламическая импульсация. Они задействованы в осознанном получении учёным удовольствия, например от чтения данного выступления. И наконец, нейроны круглой формы – получение удовольствия без помощи сознания. В прилегающем ядре более 90% нейронов это ГАМК нейроны, то есть тормозящие. Это следствие того, что в обычной ситуации плюс нейроны должны быть заторможены и также того, что с помощью ГАМК проекционных нейронов осуществляется торможение или активация двигательных или вегетативных нейронов, направленных, как на получение ощущения удовольствия, так и на прерывание отрицательных двигательных и вегетативных воздействий. Наличие в прилежащем ядре волокон связи с медиодорсальным ядром таламуса означает участие нейронов удовольствия, в том числе, фиксации в префронтальной коре блоков памяти с положительными эмоциями, то есть в комбинировании разных компонентов. Гиппокамп в прилегающем ядре участвует через свой универсальный выключатель, то есть через субикулум. Как было сказано ранее, в эволюционном развитии появление раздельнополых организмов дало большой скачок в увеличении морфологической сложности организмов за счёт активного перемешивания геномов, то есть должен был сложиться механизм, заставляющий раздельнополых особей стремиться найти друг друга. Основой такого механизма и стали нейроны удовольствия. Во всём многообразии такой механизм стал функционировать с появлением позвоночных животных, или что то же самое пейсмекерных нейронов. Принцип действия нейронов удовольствия похож на действие болевых нейронов. Если там прохождение потенциала действия по болевому нейрону вызывает разбалансировку механизма аксонного транспорта, то здесь, наоборот, прохождение потенциала действия должно приводить или к остановке аксонного транспорта, или генетически закреплённый механизм скольжения компонентов аксонного транспорта внутри нейрона. Отсюда следует весьма парадоксальный вывод: обычный нейрон с обычным аксонным транспортом должен вызывать во время потенциала действия легкое отрицательное последействие, а так как организм это не ощущает, то он появляется на свет с нейтральным восприятием этого отрицательного последействия. Для организма это начало отсчёта: в одну сторону боль, в другую удовольствие или полное отсутствие легкого отрицательного последействия, то есть нынешней нормы.
Миндалина
После появления нейронов с самоинициируемой произвольной импульсацией, то есть с появлением позвоночных животных, когда особи не надо ждать, чтобы тебя дёрнули за хвост для каких-либо действий, появилась возможность начать строить морфологически сложный организм с постоянно работающими органами и механизмами. Сложный организм стал быстрее добывать себе пищу. Возникло свободное время между периодами приёма пищи за счёт избыточного питания. Начали образовываться связи между плюс нейронами и различными этапами добычи приёма, переработки пищи, как самого важного для организма этапа. Рядом с прилежащим ядром начали образовываться кортико-медиальные ядра миндалины с различными этапами обработки пищевой деятельности. Одновременно возникали связи между болевыми нейронами и различной деятельностью организмов, которые приобретали для организма значение опасности. Причём всё это формировалось, как бессознательные процессы. И если в прилежащем ядре в одном месте находятся нейроны активные, как на бессознательном уровне, так и на сознательном, то в миндалине участие сознания привело к появлению базолатеральной группы. И так как у приматов, а особенно у человека большая часть времени тратится на борьбу за выживание и в первую очередь между соплеменниками, сознательная часть миндалины – базолатеральная часть резко разрастается. Это приводит к тому, что общая деятельность миндалины становится резко отрицательной. Можно сказать, что миндалина это орган, переводящий всю жизнедеятельность организма на положительно-отрицательное восприятие. Когда положительное восприятие, то импульсируют нейроны удовольствия прилежащего ядра, когда отрицательное, то болевые нейроны. Выход миндалины, как и любого самостоятельного ядра, будет идти по двум направлениям: различного рода двигательная активность и различного рода вегетативная активность, то есть направленная наружу и направленная внутрь. Остается сказать по этому пункту, что все процессы построения миндалины начались на основе обоняния.
Обоняние
Обонятельная система человека, как и вкусовая устроена достаточно просто. Центральный мотив обоняния соединения пространственной конфигурации пахучей молекулы в обонятельных рецепторах, проведение возбуждения в клубочек обонятельной луковицы с наложением ощущения в том же клубочке активации того или иного количества нейронов четырех типов, расположенных в четырех переднезадних полосах или зонах экспрессии. Четыре типа нейронов – это болевые, холодовые, тепловые и удовольствия, которые, как и во вкусовой системе, дают ощущение горького, кислого, соленого, сладкого. Причём расположение зон чувствительности на языке дает подсказу эволюционного времени возникновения данного ощущения. В эмбриогенезе по мере образования языка видно, что сначала появились нейроны боли, то есть зона горького вкуса, потом холодовые нейроны – зона кислого вкуса, потом тепловые нейроны – зона соленого вкуса, потом нейроны удовольствия – зона сладкого вкуса. Как уже было сказано, что сознание строилось на обонянии, как на самом сложном виде чувствительности. Роль таламуса исполняют митральные клетки за счёт постоянства периодического возбуждения. Для гиппокампа была взята конструкция с обонятельной луковицей, когда зубчатая фасция гиппокампа тормозит зону CA 3. В обонятельной луковице зернистый слой тормозит слой митральных клеток. Дендритное дерево гранулярных клеток зубчатой фасции гиппокампа, которая образует униполярный конус апикальных дендритов такое же, как и у клеток зерен в обонятельной луковице. Септум, как инициатор осознанных процессов начал строиться на торможении обонятельного возбуждения. Вентральная тегментальная область участвует в общей активации центральных сознательных процессов, как в обонятельной системе. Обонятельный бугорок выполняет функции похожие на функции медиодорсального ядра таламуса. Ограда начала формироваться на первичной обонятельной импульсации, беря информацию из обонятельной луковицы, обрабатывая её, и, посылая её, в самые первичные миниколонки в обонятельной коре. Остается только добавить, что осознанное обонятельное ощущение у человека будет исходить из области переднего конца гиппокампальной извилины, из поля 34. Кора в этой области, как обычно, для зон восприятия имеет не шести, а трёхслойное строение.
Островок
Островковая кора – это область новой коры, полностью построенная на импульсации болевых нейронов. Также как и в других чувствительных областях коры вторичная зона островковой коры передняя построена с помощью ограды. Островок причастен, как ко всем видам отрицательных эмоций: страха, гнева, тоски, ипохондрии, безнадежности, отчаяния и так далее, так и к различной дестабилизации внутренних вегетативных процессов: изжоги, поноса, рвоты и так далее. Островок будет формироваться пропорционально общему количеству первичных болевых нейронов. Это говорит о том, что в первичной коре островка больше будет болевых нейронов. И у более крупных животных с одинаковой морфологической сложностью более вероятно большее развитие передней инсулярной коры. Например, у гориллы более развита фронтоинсулярная кора, чем у карликовых шимпанзе, хотя последние гораздо ближе к человеку по морфологической сложности. Это видно по наличию веретенообразных нейронов постоянно проводящих возбуждение в передней островковой коре. Именно островок является основным поставщиком возбуждения от болевых нейронов для отрицательных программ в миндалине, как и прилежащее ядро для положительных программ. Островок посылает афференты во все места, где требуется какого-либо рода отрицательная информация. Для памяти кратковременной – в поле 10 префронтальной коры, для долговременной памяти – в поле 9, для осознания – в гиппокамп, для формирования разного рода расстройств внутреннего состояния – в гипоталамус, для ощущения вины – в медиальную префронтальную кору, для угнетения радостного возбуждения – в орбитофронтальную кору, в медиодорсальное ядро таламуса - для установления мгновенной связи с другими ощущениями, в прилежащее ядро – для торможения первичного удовольствия и многое другое.
Базальные ядра: бледный шар, скорлупа, хвостатое ядро
Для того, чтобы начать разбираться с таким достаточно сложным образованием, как базальные ядра, а это: бледный шар, скорлупа, хвостатое ядро сформируем определённые исходящие данные, а некоторые отметим ещё раз, а это: таламус периодически возбуждает те участки мозга куда приходит его волокна, каждое ядро таламуса куда бы не приходили его волокна используется для выполнения только одной функции, все нейроны таламуса используемые для выполнения одной функции находятся в одном месте таламуса. Гамкергические проекционные волокна стриатума и бледного шара не являются тормозными по определению. Наличие ГАМК медиатора в этих волокнах служит для обнуления тех возбудительных процессов, которые происходят в нейронах приемниках, а реальные процессы возбуждения или торможения определяются наличием пептидов субстанции П, динорфина, энкефалина. Энтопедункулярное ядро у простых позвоночных организмов появляется латеральнее вентрального и дорсального таламусов – аналогов таламуса и субталамуса у млекопитающих и выполняет функцию, в первую очередь, торможения возбуждающих таламических нейронов. У организмов, у которых нет торможения вентрального таламуса, а это определённые виды хрящевых рыб, например акул, будет наблюдаться постоянное движение, а чтобы организм в данный момент не двигался, то должно быть надёжно заторможено субталамическое ядро или вентральный таламус. Изначально при возникновении вентрального таламуса иннервировал все мышечные клетки. И поэтому волокна, идущие в субталамус или выходящие из него, связаны с активацией или торможением различного рода двигательных процессов. Ранее было сказано, что центром образования неокортекса является нынешний 4-й и 5-й слои, то есть слои прихода информации в кору и выхода информации из коры. Остальные слои это внутренняя переработка в коре и различного рода торможения приходящих в кору возбудительных процессов. Центрами миниколонок в неокортексе являются или модально-территориальные нейроны, например, зрительные из определённой области сетчатки, тактильные из определённых частей тела, которые расположены в первичных сенсорных и двигательных полях, или оградные нейроны, которые перерабатывают первичные модальные сигналы. Отсюда следует, что любые новые образуемые функции будут появляться, как системы приёма передачи сигналов. Эмбриологически отслежен следующий порядок возникновения ядер головного мозга. Таламус с появлением самовозбуждающихся клеток, энтопедункулярное ядро, как средство в основном торможения нейронов таламуса, кортико-медиальная часть миндалины, как место конвергенции обонятельных стимулов и различного рода вегетативных и двигательных эффектов, медиальная часть прилежащего ядра, как скопление нейронов удовольствия и связь их с обонятельно-двигательными и обонятельно-вегетативными рефлексами миндалины и наконец образование скорлупы с наружной частью бледного шара, как вариант около сознательной двигательной деятельности. Это делает возможным после образования неокортекса. Последнем в этом ряду будет появление хвостатого ядра по принципам заложенным в скорлупе и наружном бледном шаре. Исходя из изложенного и зная, что хвостатое ядро отличается от скорлупы наличием головки и формы опоясывающего разомкнутого кольца несложно определиться с предназначением и работы базальных ядер. Скорлупа вместе с бледным шаром, таламусом, субталамусом, компактной чёрной субстанцией и педункуло-понтийным ядром осуществляют около сознательную деятельность или по-другому вместе с сознательной, то есть сосознание. Например, гепард, захватывая клыками шею антилопы, совершает определённый набор движения головой и туловищем. Этот набор движений активируется из дополнительной моторной области, из которой вызывается и ещё целый ряд двигательных программ по различным аспектам жизнедеятельности. Главное, что их объединяет это то, что функционируют они только с сознанием и это ни осознанная деятельность, ни рефлекторная, например одергивание руки после укола. Само создание скорлупы шло, как развитие различного рода осознанного движения для получения удовольствия от приёма пищи с использованием медиальной части прилежащего ядра. И поэтому в основе скорлупы лежат так называемые стриосомы, то есть нейроны полностью идентичные нейронам первоначальных полей некоортекса нынешних 4-х и 5-х слоев, на которые конвергируют аксоны миндалины с положительным подкреплением. Все это базировалось на постоянном возбуждении таламических нейронов средней линии таламуса во время бодрствования. Вообще-то средняя линия таламуса является скоплением одних из самых первых нейронов и по степени дифференцировки в эмбриологическом развитии, и по месту расположения. Нейроны средней линии таламуса являются основой сознания и всех первоначальных процессов, связанных с сознанием в скорлупе и в хвостатом ядре. Так как по законам формирования мозга новая сложная функция образуется, как совокупность приёма и передачи информации, вспомним 4-ые и 5-ые слои неокортекса, то стриосомы скорлупы являются местом приёма информации необходимой для формирования полноценных двигательных программ, куда приходит информация из новой коры и 5-го слоя моторной коры, а также 4-го и 5-го слоя сенсорной коры. Таким образом, скорлупа является аналогом 4-го слоя неокортекса, следовательно аналогом 5-го слоя неокортекса будет внешний сегмент бледного шара. Каждый проекционный гамкергический нейрон скорлупы является полным аналогом центрального нейрона миниколонки неокортекса, является полным аналогом проекционного нейрона наружного бледного шара, является полным аналогом проекционного гамкергического нейрона внутреннего бледного шара. Если от нейронов дополнительной моторной области коры начинаются большие сложные программы движения, то от проекционных гамкергических нейронов скорлупы начинаются или продолжаются более простые блоки движений, которые идут до внешнего бледного шара. Далее импульсация идёт или к таламусу, или субталамусу для включения или выключения соответствующих возбуждающих таламических нейронов, а также импульсы идут назад в скорлупу для инициации следующего нейрона, иннервирующего через ряд переключений следующую мышцу. Все нейроны одного блока движений в скорлупе располагаются на одном горизонтальном уровне. Каудально в скорлупе будут располагаться блоки самых первых двигательных программ у этого эволюционного ствола организмов, а рострально – самые последние. Сами блоки нейронов составляются с помощью медиодорсального ядра таламуса, которое функционально связывает разные проекционные нейроны в сеть и с помощью центромедиального интраламинарного ядра таламуса, которое увеличивает длину волокон. С латеральной стороны скорлупы будут располагаться нейроны аналогичные первичным моторным и сенсорным полям неокортекса. Соответственно, медиальнее по направлению к таламусу в скорлупе будут располагаться аналогичные нейроны уже задействованные в составлении блоков различных двигательных программ. С самой медиальной стороны в скорлупе располагаются нейроны начала двигательных блоков. Скорлупа, как и бледный шар, является полосатым телом. Полосы образованы миелиновыми волокнами проекционных гамкергических нейронов. Они очень похожи структурно на полоски Белларже в неокортексе. Разница только в том, что там они состоят из безмиелиновых волокон, а здесь волокна в миелиновой оболочке. Это говорит, как мы уже знаем о том, что поток импульсации по стриарным проекционным волокнам гораздо больше, чем по аксонам в коре. Получаются полосы следующим образом. В норме волокна будут располагаться вокруг скопления нейронов. Первоначально скорлупу образуют нейроны 4-ых и 5-ых слоев неокортекса. Потом развитие коры привело к появлению 3-го слоя и 2-го слоя, соответственно. Разделив число полос скорлупы на три части, найдём количество морфофункциональных объединений составляющих тело организма. Объединение скорлупы и бледного шара является единственным около осознанном центром скопления различного рода двигательных программ. Поэтому каждое новое функциональное ядро, например, базолатеральное ядро миндалины или новые нейроны неокортекса, будут являться инициаторами образования новых двигательных программ в скорлупе и бледном шаре. Каждая новая программа будет располагаться ростральнее. Морфофункциональные механизмы стриатума и паллидума очень сильно отличаются от неокортекса. Если в новой коре всё возбуждение дают ядра таламуса, то в стриатуме аналогичные функции выполняют только ядра средней линии таламуса. Остальные ядра таламуса выполняют функции развития стриатума или связаны с выполнением определённой работы стриатума, но главная функция такого возбуждения в конечном нейроне, которая будет достаточна для его возбуждения в определённый момент. И все стриосомы через ядра средней линии запитаны этим возбуждением, тогда как матрикс лишен этого. Матрикс получает возбуждение за счёт ацетилхолиновых нейронов, которые, получая возбуждение от стриосом, передают его по всему объёму стриатума. И чем сильнее будет активация стриосом, тем сильнее будет возбуждение окружающего его матрикса. Стриосомы получают большую активацию от миндалины, и от орбитофронтальной коры, и от медиофронтальной коры, то есть от эмоциональных центров. И чем больше будет эмоциональный всплеск, тем большее возбуждение пойдёт от стриосом в компактную чёрную субстанцию, тем сильнее будут активированы дофамином все его получатели, и соответственно, в зависимости от вида дофаминовых рецепторов или будет увеличено возбуждение, или будет увеличено торможение соответствующих нейронов. Это может привести и к такому следствию, когда в результате эмоционального стресса через стриосомы скорлупы будет активированы двигательные волокна в несколько раз сильнее, чем в норме. Различают два типа проекционных нейронов. Более многочисленные нейроны с1 среднего размера это сенсорные нейроны, которые дают многочисленные коллатерали, а более крупные, но в меньшем количестве нейроны с2 это двигательные. В стриатуме реализуется система быстрого образования новых нейронов и новых связей между нейронами, в отличие от неокортекса, через активацию соматостатин содержащие без шипиковых нейронов. Если скорлупа функционирует в системе сосознания, то хвостатое ядро, которое появляется эволюционно позднее, является центром подсознания или того, что у нас именуется кратковременной памятью. Наличие головки у хвостатого ядра в отличие от скорлупы, которая активируется от неокортекса и функционирует с постоянным окружении с ним говорит о том, что оно может активироваться независимо от нынешней сознательной деятельности. Например, человек идёт и разговаривает, разговаривает – это сознание, а ходьба осуществляется через активацию головок хвостатого ядра – это подсознание. Оно происходит отдельно от сознания и принципиально отличается от сосознания. Переключение с подсознания на сознание осуществляется через педункуло-понтийное ядро. Работает хвостатое ядро следующим образом. Нейроны неокортекса в результате различной осознанной деятельности одновременно активируют аналогичные нейроны в хвостатом ядре, дальше информация с проекционных гамкергических нейронов идёт во внутренний бледный шар, потом на таламическое вентральное переднее ядро VA и далее в поле 10 префронтальной коры, там она будет храниться некоторое время до получаса, после чего будет стерта следующей информацией. У нейронов вентрального переднего ядра соматопическая организация. Нейроны ядра обычно подтормаживаются, но после поступления активированного сигнала поддерживают его на определённой части нейронов поля 10. Записи этих сигналов в поле 10 происходят с помощью медиодорсального ядра таламуса, продолжительность импульсации нейронов префронтальной коры зависит от числа нейронов в поле 10 и от величины активации реализуемой через стриосомы хвостатого ядра, как у стриосом скорлупы. И поэтому продолжительность кратковременной памяти, а это оно и есть, пропорционально эмоциональной значимости стимула. Доставание из кратковременной памяти будет происходить следующим образом: передняя поясная кора, поле 10 префронтальной коры, головка хвостатого ядра и далее в неокортекс в те зоны, где находятся зеркальные нейроны и далее восприятие, как уже было рассказано. Основным базовым элементом подсознания является переднее вентральное ядро VA таламуса, то есть куда приходит его волокна, те части мозга принимают участие в процессах подсознания, а вентральное латеральное ядро VL – это основное двигательное ядро без участия сознания. Хвостатое ядро своим круговым расположением представляет собой единственный сочный пример, когда эволюционно поздняя возникающая структура развивается в каудальном направлении вопреки законам развития головного мозга гласящим, что новые возникающие ядра мозга формируются только с ростральной или латеральной стороны от уже существующих. Это происходит потому, что развиваться хвостатое ядро может только в направлении волокон выходящих из головки хвостатого ядра. Сначала рост хвостатого ядра происходит рострально под неокортексом, а потом хвост заворачивается в виде кольца. Определяющим здесь будет возможность конца хвоста растолкать уже сформированную структуру, невозможно пройти через костную ткань мозга и через уже работающие ядерные образования, слишком много взаимопроникающих волокон между клетками ядра, как в клубке. Остается только проходить через пучки проводящих волокон, которые при этом будут слегка удлиняться. Так как субталамическое ядро является правопреемником вентрального таламуса, который формировался как постоянный возбуждающий поток на мышечные клетки, то понятно, что все волокна, проходящие или уходящие от субталамического ядра имеют отношение к различного рода двигательной активности. Внутренняя часть бледного шара вся имеет отношение к возбуждению или торможению движения, а наружная часть бледного шара связана с двигательной активностью в русле волокон связанных с субталамическим ядром. Так как уже известно, что хвостатое ядро является центром подсознания, то понятен и механизм этого процесса. Какие-либо события, происходящие в фокусе внимания человека активируют нейроны неокортекса, принимающие участие в восприятии этого. Одновременно с этим с этих нейронов происходит возбуждение на аналогичные нейроны хвостатого ядра. Потом в зависимости от эмоциональной нагрузки, проведённой через стриосомы более сильный или более слабый поток импульсов идёт на ретикулярную часть чёрной субстанции, которая активирует или тормозит аналогичные нейроны вентрального переднего ядра таламуса, из которого импульсы достигают поля 10 префронтальной коры. Связанная между собой с помощью медиодорсального ядра таламуса в хвостатом ядре информация доставляется с помощью вентрального переднего ядра, а развешивается на нейронах поля 10 коры. Чем больше нейронов поля 10, тем на большем количестве нейронов будет развешена эта информация. Увеличение времени нахождения информации в памяти или подсознания, что более правильно, будет способствовать увеличенная дофаминергическая и ацетилхолинергическая импульсация. Несложно представить, что организму надо, чтобы все выходные нейроны вентрального переднего ядра таламуса были бы постоянно подторможены, а активироваться они будут при осознаваемых процессах в неокортексе. В заключение этого раздела скажем, что дофаминергические системы а8 ретроробральная зона, а9 компактная чёрная субстанция, а10 вентральная тегментальная область появились в эволюции совсем в обратно порядке – сначала а10, потом а9 и наконец а8. Вентральная тегментальная область а10 – это доведение возбуждения до 100 процентов от требуемых для инициации потенциалов действия в центрах участвующих в первичных сознательных процессах.
Сон
Следующая тема, которую хотелось бы затронуть это сон. В этой теме наиболее ярко проявляется полная невозможность познания головного мозга без привлечения здравого смысла, а только в результате разнообразных экспериментов с использованием различного вида оборудования, в том числе и очень дорогостоящего. Это получается потому, что сон представляет собой глобальную функцию организма и задействовано в нем в той или степени большая часть всех нейронов головного мозга. Поэтому, какой нейрон не возьми он будет либо ухудшать, либо улучшать сон. Здравый смысл говорит, что когда работает какой-либо механизм, то всегда происходит расходование элементов требуемых для его работы. Для того чтобы определить, что это за элементы посмотрим на самую показательную в этом отношении часть сна – первую половину сна. Понятно, что после бодрствования наступает переходный период – часть сна стадия 1 и стадия 2, как результат включения эволюционного выработанного механизма активации сна и деактивации бодрствования с помощью преоптической передней области гипоталамуса. А дальше наступает 3-и и 4-е стадии сна, которые и являются в нашем случае лакмусовой бумажкой в познание. В данное время в ЭЭГ неокортекса наблюдается в большинстве своем дельта-ритм – это и есть показатель реального состояния организма после длительного бодрствования. Рассматривая какие обстоятельства приводят к появлению дельта-ритма в коре обнаруживаем, что к этому приводят за исключением различного рода расстройств и травм у полностью здорового человека уменьшение содержания кислорода в крови и уменьшение содержания сахара в крови, то есть тех элементов, которые нужны для образования аденозинтрифосфата – основного энергетического источника для функционирования любого нейрона. Поэтому констатируем, что сон, который присущ всем позвоночным – это процесс восстановления работоспособности организма. Дельта-ритм во сне и не только проявляется, когда у авторитмичных нейронов из-за недостатка энергии происходит сбой работы и вместо равномерного запуска потенциалов действия происходит группирование их в пачки, но у сна есть такая разновидность, как быстроволновый сон или сон с быстрым движением глаз – бдг-сон, который портит жизнь всем нейрофизиологам и не даёт возможность разобраться во сне. Разбираться с быстрым сном будем, опять же, с помощью здравого смысла, а ключом здесь будет период в 90 минут, с которым в среднем происходят циклы бдг-сна. Если покопаться в физиологии человека, то можно найти достаточное количество циклов продолжительностью в 90 минут. Это периодическая моторная деятельность желудочно-кишечного тракта, когда с периодом 90 минут происходит увеличение, а потом уменьшение сократительной активности конкретного участка кишечника. Далее первый эпизод рем сна наступает через порядка 90 минут. Далее при резком снижении концентрации глюкозы в кровяной плазме – гипогликемии – в результате работы нервной эндокринных систем возвращается к норме концентрация глюкозы в крови с периодом до, опять же, тех же 90 минут. Ну и отмеченная психологами периодичность улучшения работоспособности людей с циклом тех же самых 90 минут, ну и многие другие циклы. Чтобы разобраться в быстроволновом сне вспомним, что он есть только у птиц немного и у млекопитающих много. Млекопитающие от других организмов отличаются резким увеличением морфологической сложности, в следствии чего развиваются системы на чём держится это увеличение сложности. Это система сердечнососудистая почечная, то есть всё, что связано с циркуляцией крови. Важным фактором этой системы является вазопрессин, влияющий и сам и через другие гормоны на автоматическую регуляцию крови и межклеточного пространства. В результате действия или бездействия вазопрессина изменяется соотношение натрия, кальция, калия, хлора в межклеточном пространстве. Это приводит к изменению возбудимости нейронов, а это уже у низкопороговых нейронов может привести к возникновению потенциалов действия в результате одиночного стимула. Если во время бодрствования такое срабатывание устойчиво подавляется постоянным подтормаживанием систем, то во время сна это выливается в бдг-сон. И поэтому быстроволновый сон начинается в структурных формированиях самых низкопороговых нейронов: миндалине и гипоталамусе. Из-за того, что вовлекаются миндалины почти все сновидения имеют эмоциональную окраску. Сами по себе сновидения являются следствием постоянного расположения в энторинальной коре временного хранения нейронов, в которых закодированы информационные блоки продолжительностью до 12 секунд, которые потом перезаписываются в те или иные участки неокортекса: лобного, височного, теменного. Если во время медленноволнового сна в самом начале сна сновидения являются результатом планового официального перезаписывания, то в бдг-сне они будут следствием срабатывания самых низкопороговых нейронов, которые вытаскивают самые низкопороговые блоки информации энторинальной коры, её глубоких слоев. И поэтому человек в своих снах очень часто видит одни и те же события. Все сновидения строятся только из ранее записанных блоков информации, хранящихся в энторинальной коре. Все структуры мозга ответственные за приём новой информации во время сна отключаются. Какая-либо новая информация, например, льющаяся вода во сне будет оформляться в образе дождя из ранее записанного дождевого блока информации. Соединяться это будет в большом гипппокампе или маленьких гиппокампах энторинальной коры, посредством связей с собственным я в медиофронтальной коре. Таким образом, сновидения происходящие в быстроволновом сне являются своеобразным плаваньем по волнам памяти, в структурах осознавания. И это такой побочный продукт такого эволюционного развития, когда сначала создается структура осознавания, потом механизм перезаписи информации, а потом при морфологическом усложнении возникают предпосылки самопроизвольного процесса перезаписи без окончательной новой записи. Возвращаясь к 90 минутному циклу, можно сказать, что это время обусловлено продолжительностью системы деактивации гормонального воздействия, происходящее в сердечно-сосудистой почечной системе. На цифру 90 минут в первую очередь влияет величина общего количества крови в системе. У новорожденных малышей, у которых объём крови совсем небольшой величина быстроволнового сна сопоставима с медленноволновой. И в заключение этого раздела можно сказать, что с появлением бдг-фазы сна в организме начали происходить изменения направленные на борьбу со следствием того, что происходит в эту фазу. В этом состоит изюминка развития мозга, когда не убирается побочный продукт развития мозга, а нивелируются последствия его действия.
Развитие
Рассмотрим некоторые аспекты развития мозга у взрослого организма. Для того, чтобы нормально функционировать на этой Земле любое животное должно адекватно реагировать на различные изменения в окружающем мире, то есть должны устанавливаться новые нейронные связи, должны появляться новые нейроны. Это будет выражаться в занесении в память, в образовании автоматизма движений, да и в изменениях в самом организме. Если в эмбриогенезе всё понятно развитие идёт по заранее записанной программе, то постэмбриогенезе после рождения и особенно после взросления современная наука начисто отрицает возможность и даже не рассматривает механизмы по образованию новых связей и новых нейронов у животных и у человека тоже, а они есть. В первую очередь это цереброспинальная жидкость, которая содержит не только отходы от работы нейронов, но и вещества необходимые для текущей работы нейронов, например, глюкозу, а также вещества для развития и образования нейронов. Находится цереброспинальная жидкость в желудочках мозга и спинномозговом канале. Расположение нейронов центрального серого вещества вокруг спинномозгового канала и даже не в один слой показывает, что к большому количеству нейронов разной медиаторной принадлежности требуется связь со спинномозговой жидкостью, и соответственно, практически каждое образование головного и спинного мозга будет посылать волокна к месту нахождения цереброспинальной жидкости, а особенно ярко это демонстрируется в формах боковых желудочках головного мозга, которое полностью повторяет форму полушарий головного мозга. Понятно, что не полушария повторяют форму желудочков с этой жидкостью. И чем больше полушария, тем больше боковые желудочки, которые будут расти в следствии увеличения вводимых в желудочки разнообразных веществ, соответственно, увеличение давления внутри них. Исследователям не сложно будет найти соответствие количества нейронов неокортекса и объёмов спинномозговой жидкости в желудочках. Эволюция может пойти и другими путями обеспечения нейронов необходимыми элементами. Если к какому-либо образованию головного мозга требуется массовое появление связей и новых нейронов, то для своей деятельности происходит размещение внутри этого образования целой группы нейронов. Основной целью которых является обеспечение при проведении по ним потенциалов действия окружающих нейронов веществами для развития и функционирования. Например, это класс вставочных гамкергических нейронов в скорлупе и хвостатом ядре. Наличие в них молекул ГАМК, то есть гамма-аминомасляной кислоты означает, что для того, чтобы в проекционных нейронах начались активные процессы развития надо остановить текущую деятельность конкретного нейрона, что и происходит с помощью тормозного медиатора ГАМК, но первым в ряду участников обеспечения развития нейронов является интерпедункулярное ядро, которое ранним созреванием в онтогенезе доказывает это. И несложно сделать вывод, что там куда приходят волокна из этого ядра постоянно образуются новые связи и нейроны. У межножкового ядра большое количество разнообразных связей. Самые многочисленные из них это связи с медиальным ядром поводков, причём обнаружена чёткая топологическая закономерность в организации этих связей. Вообще-то для головного мозга характерны чёткие топологические закономерности в связях между различными участками и перечень этих связей будет всегда один и тот же. Наиболее отчетливо он выявляется в неокортексе. Каждые элементы этих связей это или модально-территориальный сигнал, или дифференцированно-оградный сигнал, которые являются, в свою очередь, центрами миниколонок неокортекса сзади центральной борозды. И эти нейроны, несущие разные сигналы надо в разных частях мозга по-разному обработать. Исходя из положения нейрофизиологических законов, которые гласят, что выходные действия любого ядра будут всегда однотипными, можно констатировать, что интерпедункулярное ядро в этом плане в мозге стоит особняком. Оно свои действия распространяет, как на нейроны других ядер, так и на спинномозговую жидкость. И объединяя эти два направления, а это напомню: ядро в основную функцию которого входят образование веществ для развития нейронов и доставка их к месту потребления, а также нахождение в этом ядре полного перечня чувствительных и переработанных в ограде чувствительных клеток. Приходим к выводу, что в головном мозге, да и в спинном тоже, существует очень древняя система выборочного развития определённых клеток. Функционировать она будет следующим образом. Если есть прямые эфферентные связи между межножковым ядром и ядром получения, допустим, передним ядро мамиллярных тел, то здесь всё понятно, а когда нет прямых связей, то здесь на помощь приходит спинномозговая жидкость. Нейроны межножковых ядер выбрасывают гранулы в спинномозговую жидкость, а нейроны, например, неокортекса, которым предназначены они, с помощью маркеров вылавливают их из этой жидкости. Дальше активное вещество с помощью аксонного транспорта поступает в тело клетки и инициирует различные физиологические процессы. Информацию нейроны межножкового ядра получают от поводковых ядер, а поводковые ядра от медиодорсального ядра таламуса, которое, напомню, является местом формирования новых связей. Особое место в этой схеме играют поводковые ядра. Разберём их предназначение. Из законов нейрофизиологии известно, что в любом функциональном блоке сначала появляется скопление нейронов, выполняющие основную функцию, а потом латеральнее, просто больше для них нет места, появляются группы нейронов обслуживающих основные нейроны или каким-либо образом видоизменяющие их действие. Так формировались и поводковые ядра. Медиальное поводковое ядро образовалось, как место вычленения тех чувствительных и оградных нейронов, которые в данный момент участвуют в образовании новых осознанных связей, а латеральное поводковое ядро формировалось, как место сосредоточения обслуживающих нейронов, например, связанных с центральным серым веществом вокруг сильвиева водопровода. И только значительно позднее, когда медиальное поводковое ядро было образовано, и когда начала возникать стриопаллидарная система, а это, напомню, подсознательная двигательная и когнитивная деятельность, то латеральнее медиального ядра начали появляться нейроны, участвующие в образовании новых уже подсознательных связей. И получилось, что в одном ядре – в латеральном ядре поводков – сплелись две функции. Функция обслуживания и корректировки медиального ядра поводков, и функция развития подсознательных процессов в стриопаллидарном комплексе. Причём волокна, исполняющие первую функцию, будут проходить ядро основного получателя – интерпедункулярное – без переключений, а волокна второй функции будут в основном предназначены этому основному получателю. В общих словах действие системы образования новых связей и нейронов, например, в теменной ассоциативной коре выглядит следующим образом. Во всех участках мозга, где образуются новые связи, которые расширяют возможности уже существующих, находятся принимающие волокна медиодорсального ядра таламуса. Во время инициируемого медиальным септальным ядром тета-ритма, когда в гиппокампе происходит активный процесс осознавания текущей ситуации, в медиодорсальном ядре таламуса проходит связывание всей информации в момент времени. Причём это будет возможно только во время тета-ритма гиппокампа. Связанная во время информация по поводковому пучку волокон на топологически однотипные нейроны поступает в медиальное поводковое ядро, откуда она направляется в межножковое ядро на такие же однотипные клетки, но уже с секреторной активностью. Дальше произведенный секрет выводится в спинномозговую жидкость, потом этот секрет вылавливается из жидкости нейронами теменной зоны, участвующими в проведении потенциалов действия во время тета-ритма медиального септума. Понятно, что подведение к клеткам активных веществ по аксонному транспорту и увеличение импульсации по интраламинарным нейронам должно быть согласовано, ведь интраламинарные нейроны выполняют функцию увеличения длины волокон нейронов. Следующее образование, которое мы рассмотрим в этом ряду это дорсальное тегментальное ядро или ядро Гуденна. Даже простой анализ связей этого ядра в первую очередь с моторными ядрами понтомедуллярного отдела и вегетативными ядрами понтомедуллярного отдела показывают, что это ядро отвечает за соединение двигательной активности в первую очередь мышц головы и шеи с вегетативными реакциями в любых ситуациях. Например, при испуге комплексов движений мышц лица, головы, тела будут соответствовать конкретные изменения в физиологических параметров: частоты сердечных сокращений, артериального давления, активации потовых желез и так далее. Ядро Гуденна, как и многие образования головного мозга принимает участие в двух направлениях своей деятельности: участие в записи в память конкретных событий, и участие в развитии и увеличении амплитуды совместно двигательной и вегетативной активности. В этих процессах принимает участие мамиллярные ядра гипоталамуса и интерпедункулярное ядро. Зная их характеристики, думаю, что любой исследователь определит какое участие с каким ядром связано и без чей-либо помощи. Последним в этом ряду образований и определённым образом связанным с предыдущими ядрами является эпифиз. Эпифиз это железа, функции которой становятся понятны при сравнении организмов с эпифизом и без него. Например, человек, у которого есть эта железа и китообразные, у которых нет её. Так как наиболее активная деятельность эпифиза отмечена на начальных этапах развития, то несложно догадаться, что человек в раннем развитии от китов отличается не пропорциональным развитием, когда как у китов развитие идёт строго пропорционально в размерах тела. У человека при рождении голова по высоте составляет приблизительно 25 процентов от общей длины тела, а у взрослого около 12 процентов. Это осуществляется за счёт того, что определённые клетки головы, причём не только нейроны головного мозга или клетки черепа, затормаживаются в своём развитии, в своём делении. Этому способствует гормон эпифиза мелатонин, который имеет небольшие размеры, высоко липофилен и проникает через клеточные мембраны и ядерные мембраны, выполняет функции активации работы ДНК клетки, а это списывание с ДНК РНК и потом образование белка с РНК. Причём активироваться будет все клетки организма. Время активации клеток будет пропорционально входящему мелатонину. Другая функция мелатонина заключается в связывании с рецептором мелатонина на мембране клетки и через посредников блокирование процесса деления клетки, что делает невозможным развитие данного органа или ядра мозга. Эпифиз в эмбриогенезе организма закладывается такого размера, такого количества внутренних клеток – пинеалоцитов – в целлюлярном эпифизе, которого будет достаточно для обеспечения своей главной функции – дифференцированного развития организма, которая и осуществляется в течение жизнедеятельности организма до момента полового созревания. Половое созревание у человека обозначает время окончательного построения по записанным генетическим программам. В этот момент в несколько раз падает производительность эпифиза, который становится трабекулярным, при котором участки разросшейся стромы еще не полностью изолируют друг от друга участки паренхимы. Половое созревание означает, что организм полностью сформирован, но эпифиз продолжает функционировать, хоть и в усеченном варианте, потому что природой не вырабатывается система уничтожения органа, на основе которого строится организм и на основе деятельности которого завязаны функции многих органов. Если допустить, что после окончательного формирования организмом эпифиз бы разлагался, то это фиксировалось бы с помощью генов в ДНК. Это привело бы к тому, что в эмбриогенезе следующего поколения эпифиз начал бы деградировать раньше, чем начал функционировать, а это привело бы к прекращению развития данной эволюционной линии. Поэтому ограниченно работающий эпифиз по-прежнему будет выбрасывать мелатонин в кровь и спинномозговую жидкость. Сама по себе система эпифиза достаточно оригинальным способом решает вечную дилемму эволюции, как построить из одной оплодотворенной яйцеклетки большой сложный морфологический организм. Во время эволюционного развития на планете появились и другие варианты. Самым известным из них является метаморфоз, а именно глубокое преобразование строения организма, в процессе которого личинка превращается во взрослую особь. В основе метаморфоза лежит невозможность обеспечения полностью генетически законченного развития организма без привлечения дополнительного количества питательных веществ. Это поступление питательных веществ решается с помощью развития эмбриональной стадии, чтобы она сама себя обеспечивала пищей, а также торможением развития взрослой стадии. Этакий своеобразный вариант работы эпифиза млекопитающих. Рассматривая эпиталамус, можно отметить, что одним из краеугольных вопросов является принцип включения, выключения развития органа, клетки, ткани. Остановимся на этом подробнее. Основной гормон эпифиза мелатонин образуется из серотонина под воздействием норадреналина. В свою очередь серотонин получается из аминокислоты и триптофана в клетках эпифиза меланоцитах, но также известно о многочисленных связях эпифиза с ядрами шва. Основным выходным гормоном нейромедиатором является серотонин, то есть находящийся в эпифизе серотонин является продуктом двух систем: самого эпифиза и ядер шва. Понятно, что здесь не идет речь о том, что просто мало серотонина в эпифизе и надо откуда-то его доставить, хотя конечным превращением серотонина в эпифизе является один и тот же мелатонин, цель использования его является разной. Если целью собственного мелатонина полученного в пропорциональном морфологической сложности организма, закладываемым и пропорционально возрасту начинающем функционировать пинеалоцитам, является дифференцированное развитие организма, то целью мелатонина изготовленного из серотонина, полученного из ядер шва, является развитие тех структур мозга, которые начинают активно функционировать независимо от времени суток. Для этого рядом с медиальным поводковым ядром формируется латеральное ядро, одной из функций которого является активация ядра шва при получении новой информации из медиодорсального ядра таламуса, а потом последовательность событий будет следующая: латеральное поводковое ядро, ядро шва, эпифиз и активация мелатонином конкретных ядер, клеток участвующих в обработке новой информации.
Полушария
Рассмотрим работу полушарий и межполушарные взаимодействия. Поверхностный анализ работы полушарий мозга сразу наталкивает нас на мысль, что правое полушарие у правосторонних людей воспринимает окружающий мир таким, каким он есть, а левое полушарие кодирует окружающий нас мир в символах. Поэтому термин доминантное полушарие уходит от нас в историю, как в прочем и другие термины в современной нейрофизиологии. Как только это становится понятно, так сразу же решаются и другие вопросы в этой области. Например, один из центральных вопросов в полушариях – это снижение активности правого полушария и правополушарные нарушения у людей сопровождаются беспричинной эйфорией, легкомысленным смехом. Ослабление функции левого полушария или его нарушения ведут к беспокойству, озабоченности, пессимистическому настроению, плачу. Эволюция на Земле идёт таким образом, что чем морфологически сложнее организм, тем больше времени в борьбе он будет проводить, как с другими видами животных, так и внутри своего вида. Венцом здесь является человек, который не борется с другими видами, зато большую часть времени тратит в борьбе за место под солнцем с себе подобными. Центральным элементом здесь является восприятие отрицательной информации и мгновенный перевод её в двигательную и вегетативную активность, через правую миндалину. Понятно, что если левое полушарие отвечает за перевод осознанной информации в систему символов, а правое полушарие воспринимает окружающий мир, то основное восприятие отрицательных событий будет приходиться на правое полушарие. Для того чтобы человек понял, что это отрицательные события в правом полушарии в фронтоинсулярной области располагается большое количество веретенообразных нейронов фон Экономо, получающих импульсацию из инсулярной области от болевых нейронов. В передней инсуле болевая афферентация обрабатывается и направляется в различные места: в миндалину, в гипоталамус, в медиофронтальную кору, в орбитофронтальную кору и так далее, где для выработки отрицательных эмоций, где для погашения положительных эмоций – прямых антагонистов. Сами по себе веретенообразные нейроны предназначены для постоянного возбуждения. Это видно по форме нейронов. И наличие в правой передней островковой коре на 30% больше нейронов фон Экономо показывает, что разница в отрицательных эмоциях с права по отношению к левой гораздо больше, а радостное поведение людей в случае отключения правого полушария объясняется снятием сильного отрицательного противодействия на миндалину, орбитофронтальную кору и другие участки мозга. Это обычная реципрокная деятельность.
Мужчина и женщина
Последней крупной темой, на которой я хотел бы остановиться в кратком эволюционном разборе нервной системы это разница в психофизиологии полов. Я не буду останавливаться в закладываемых базовых принципах в головные мозги девочек и мальчиков, которые общеизвестны, а остановлюсь только на основополагающих моментах генетически закреплённых, крупных таких моментов всего три. Первый.
На рисунке головного мозга мужчин и женщин показаны усредненные линии крупных потоков возбуждения. Слева синими линиями показаны определяющие потоки в головном мозге усредненного мужчины, а справа усредненной женщины. У мужчин эти связи фронтально-теменные, а у женщин межфронто-полушарные. Здесь есть один момент, который, естественно, исследователями не может быть понятен. Если в головном мозге присутствуют достаточно большие теменные ассоциативные зоны, то связи будут, как показаны слева между лобными зонами и теменными. Если нет теменных зон или они малы, то связи в основном будут такими, как показаны справа, то есть между лобными зонами левого и правого полушария. В данном обзоре в мои планы не входит анализ работы разных ассоциативных зон, возможно позже. А возвращаясь к ранее сказанному, можно констатировать, что это варианты строения мозга не мужчин и женщин, а людей с развитыми и не развитыми теменными ассоциативными зонами. Это такие усредненные варианты, соответственно, бывают и обратные варианты у мужчин и женщин. Это следствие сложившихся стереотипов в поведении мужчин и женщин, когда перед мужчиной стоит задача принятия верного решения для обеспечения себя и своей женщины, и это следствие того, что когда работает какая-то часть мозга, то она развивается, увеличиваясь в размерах, а теменные ассоциативные зоны – это главный показатель умственного развития человека. Второй. Чем больше размеры тела человека, тем больше болевых нейронов будет приходить в инсулярную кору через таламус из спинного мозга и тройничного нерва, тем их больше будет, при прочих равных условиях, в фронтоисулярной коре, тем больше будет, опять же при прочих равных условиях, импульсировать медиофронтальная кора – источник совестливости человека, то есть величина размеров тела косвенно увеличивает адекватность поведения человека, а мужчины в среднем крупнее женщин, это верно и для больших и маленьких женщин, как в прочем и мужчин. Третий. У женщин крупнее прилежащее ядро, и соответственно, орбитофронтальная кора, чем у мужчин. Это следствие в разнице размеров чувствительности кожи на половых органах женщин и мужчин, которые иннервируются нейронами, которые при возбуждении формируют чувства удовольствия в прилежащем ядре, а прилежащее ядро это базовая основа эмоциональности человека или по-другому: чем больше ацетилхолиновых нейронов в прилежащем ядре человека, чем больше их размеры, тем выше эмоциональность человека. Главная их функция – функционирование во время оргазма. Поэтому чем больше сексуальных отношений, тем выше эмоциональность человека, потому что постоянное проведение возбуждения по ацетилхолиновым нейронам приводит к увеличению их размеров, то есть женщина рождается с генетически заложенным механизмом повышенной эмоциональности.