Транскрипция-процесс синтеза РНК по матрице ДНК с помощью ДНК-зависимой РНК-полимеразы. Она осуществляется как у прокариот, так и у эукариот. Процессы достаточно похожи, но имеют некоторые отличия.
Общие принципы транскрипции включают:
-комплементарность- взаимное соответствие молекул биополимеров или их фрагментов, обеспечивающее образование связей между пространственно взаимодополняющими фрагментами молекул,
-антипараллельность-выражается в том, что на одном и том же конце спирали одна цепь ДНК содержит (незамещенную или замещенную) группу 5 штрих, а другая 3 штрих.
-униполярность- синтез полинуклеотидных цепей происходит всегда в направлении 5' → 3',
-ассиметричность-синтез дочерней цепи идёт только на одной цепи ДНК-матрицы, вторая при этом блокирована.
Особенности транскрипции у эукариот:
-полимеразы трех типов:1,2,3 и также РНК, используемые в митохондриях и хлоропластах,
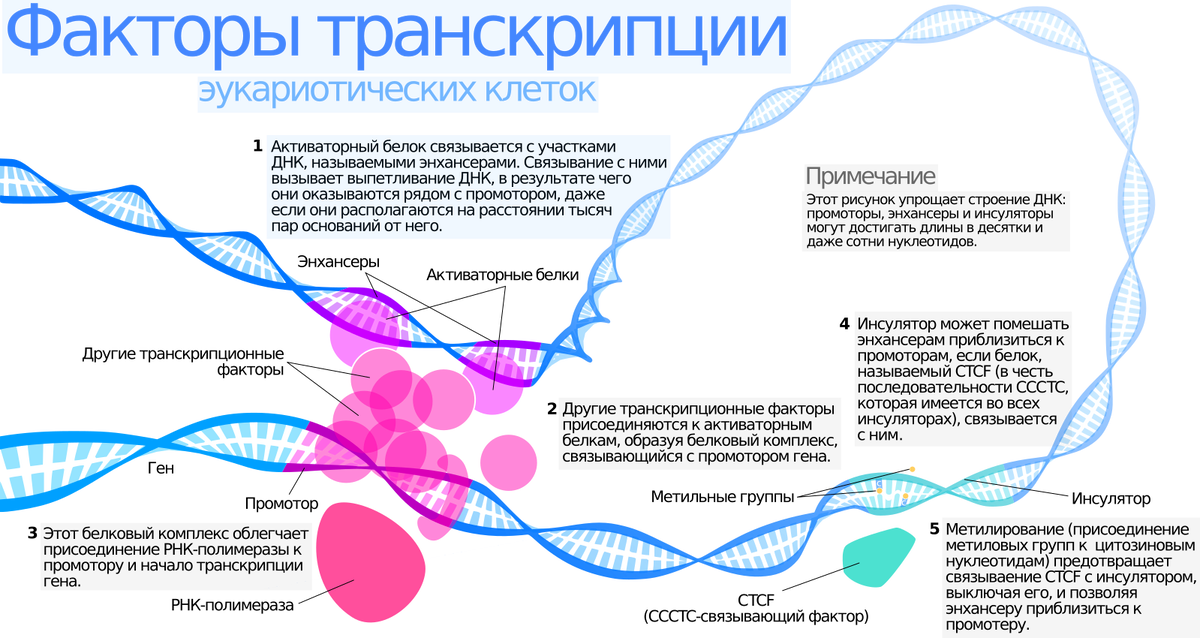
-единица транскрипции у эукариот-ген, перед геном есть последовательность-сигнал – ТАТА-бокс, которые находятся на значительном удалении от начала считывания, необходимы также белки –факторы транскрипции,
-наличие энхансеров и сайленсеров.
• Энхансеры (enchancer) — последовательности ДНК, способные связываться с факторами транскрипции, при этом увеличивая уровень транскипции гена или группы генов.
• Сайленсеры (silencer) — последовательности ДНК, с которыми связываются факторы транскрипции (белки-репрессоры), что приводит к понижению или полному подавлению транскрипции гена.
Энхансеры- несколько белков, которые взаимодействуют с последовательностью ДНК и образуются шпильки, которые выпячиваются из хромосомы и становятся более доступными для РНК-полимераз, таким образом усиливается транскрипция отдельных генов.
У эукариотов существуют различные РНК-полимеразы:
-матричная РНК синтезируется с помощью РНК-полимеразы-2,
-рибосомальная РНК синтезируется с помощью РНК-полимераз- 1 и 3,
-транспортная и малоядерная РНК синтезируется с помощью РНК-полимеразы 3.
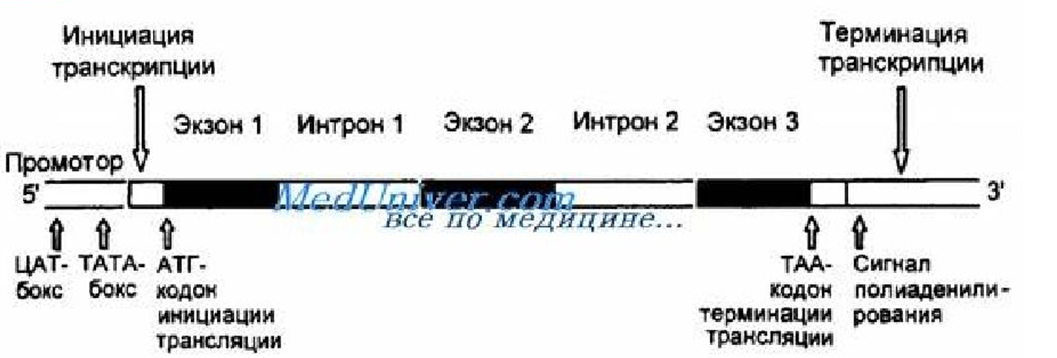
Организация генов эукариот:
-на 5 штрих конце промотор из CAAT и TATA боксов;
-для связывания промотора с первым нуклеотидом необходимы факторы транскрипции;
-считываемая последовательность начинается с кодона инициации ATG;
-кодирующая область состоит из коротких экзонов, кодирующих информацию и длинных интронов некодирующих информацию;
-заканчивается транскрипция в области терминатора;
-далее следует процессинг.
Созревание РНК или процессинг РНК (посттранскрипционные модификации РНК) — совокупность процессов , которые приводят к превращению первичного транскрипта в зрелую РНК.
В зависимости от типа РНК (матричные, рибосомные, транспортные, малые ядерные) их предшественники подвергаются разным последовательным модификациям: кэпированию, сплайсингу, полиаденилированию, метилированию и иногда редактированию.
мРНК эукариот синтезируется в виде предшественника, пре-мРНК, которые затем проходят сложное стадийное созревание — процессинг, который включает присоединение кэп-структуры к 5'-концу молекулы, присоединение нескольких десятков остатков аденина к её 3'-концу (полиаденилирование), вырезание некодирующих участков — интронов и соединение друг с другом — экзонов (сплайсинг). Соединение экзонов одной и той же пре-мРНК может проходить разными способами, приводя к образованию разных зрелых мРНК, и в конечном итоге разных вариантов белка (альтернативный сплайсинг). Только мРНК, успешно прошедшая процессинг, экспортируется из ядра в цитоплазму и вовлекается в трансляцию.
Кэпирование представляет собой присоединение к 5'-концу транскрипта 7-метилгуанозина через необычный для РНК 5'-трифосфатный мостик, а также метилирование остатков рибозы двух первых нуклеотидов. Процесс кэпирования происходит во время синтеза молекулы пре-мРНК. Кэпирование защищает 5'-конец первичного транскрипта от действия рибонуклеаз, специфически разрезающих фосфодиэфирные связи в направлении 5’→3'.
Функции кэпа и связанных с ним белков:
-участие в сплайсинге;
-участие в процессинге 3'-конца мРНК;
-экспорт мРНК из ядра;
-защита 5'-конца транскрипта от экзонуклеаз;
-участие в инициации трансляции.
Фермент поли(А)-полимераза присоединяет 3'-концу транскрипта от 100 до 200 остатков адениловой кислоты. Полиаденилирование осуществляется при наличии сигнальной последовательности 5'- AAUAAA-3' на 3'-конце транскрипта, за которой следует 5'-CA-3'. Вторая последовательность является сайтом разрезания.
После полиаденилирования мРНК подвергается сплайсингу, в ходе которого удаляются интроны (участки, которые не кодируют белки), а экзоны (участки, кодирующие белки) сшиваются и образуют единую молекулу. Сплайсинг катализируется крупным нуклеопротеидным комплексом — сплайсосомой, состоящей из белков и малых ядерных РНК. Многие пре-мРНК могут быть подвергнуты сплайсингу разными путями, при этом образуются разные зрелые мРНК, кодирующие разные последовательности аминокислот (альтернативный сплайсинг).