Делая один надрез: ДНК-никазы
Для снижения нецелевых событий редактирования Cas9 модифицировали, превратив ее в ДНК-никазу — то есть в нуклеазу, способную к расщеплению лишь одной из нитей ДНК (Cas9n).
Ну а поскольку для редактирования все же нужен двойной разрыв, Cas9n взяли в двух экземплярах, к ним же прилагая и две направляющие sgRNA. Преимущество тут в том, что отдельные разрывы восстанавливаются с высокой точностью, значительно уменьшая вероятность ошибок. В частности, одна из вариаций Cas9n — D10A — в экспериментах особо эффективно снижала индель-мутации в клеточных линиях зигот мышей.
Редактирование ДНК-оснований: Base Editing (BE) —
инновационный инструмент, в котором Cas9n соединена с ферментом дезаминазой, ответственным за прямую «конверсию» одних ДНК-оснований в другие. Например, редакторы цитозиновых оснований (cytosine base editors — CBEs) провоцируют переход цитозинов в тимины; аналогичные адениновые редакторы (ABEs) — переход аденинов в гуанины; ну а есть еще и такие, что способны триггерить одновременно оба этих перехода. И даже больше: есть редакторы, умеющие переводить цитозины как в гуанины, так и в аденины, а потом и обращать подобную конверсию вспять.
В целом, BE быстро показали свой потенциал в коррекции мутаций, ассоциированных с разными заболеваниями (далее мы еще рассмотрим это). А поскольку они не приводят к двойным разрывам ДНК, то и инделей почти не вызывают.
Однако, несмотря на немалый потенциал, BE все же не лишены недостатков и ограничений. В частности, их рабочий фермент дезаминаза имеет строго определенное «окно редактирования»: диапазон из нескольких нуклеотидов, в котором он может эффективно модифицировать нужное основание. Казалось бы: это же отлично! Редактор будет действовать в целевом диапазоне — а значит, сработает точечно. Но не все так просто: в пределах своего узкого окна BE может изменять и близлежащие к целевому участку нуклеотиды, приводя к так называемым мутациям свидетеля («bystander mutations»). Более того, несмотря на высокую специфичность, редакторы оснований могут действовать и за пределами «окна редактирования», если обнаружат участок, очень похожий на это самое окно.
Добавим, что BE направлен в основном на однонуклеотидные замены, а потому практически бесполезен, если требуется вставить или удалить более крупный ДНК-фрагмент (а значит — терапевтическая применимость тут ограничена).
«Прайм»-редактирование: Prime Editing (PE)
В сравнении с описанными выше системами редактирования PE находится на новом уровне, обеспечивая большую точность и меньшее количество ошибок. Этот инструмент может выполнять всевозможные небольшие правки и почти не приводит ни к bystander-, ни к off-target-мутациям; а индели также ему не свойственны (за исключением некоторых его вариантов, см. ниже). Причина всего этого в особом механизме действия — кстати, довольно хитроумном. Дело в том, что вместо направляющей sgRNA тут используют pegRNA, содержащую в себе также матричную последовательность для внесения в геном соответствующих целевых правок.
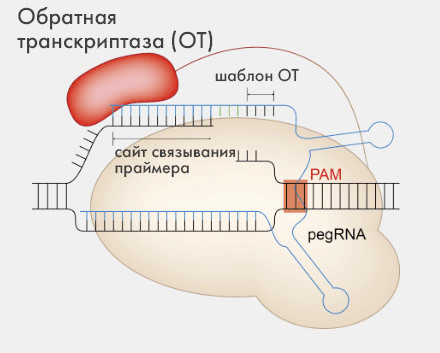
Погодите-ка, ведь для шаблона редактирования обычно берут цепочку ДНК, да и геном представлен именно этой нуклеиновой кислотой. Причем же здесь направляющая РНК? Вот в этом один из секретов! «Набор для генной редакции» в PE содержит позаимствованную у вирусов обратную транскриптазу, а фермент этот, как известно, умеет переписывать генетическую информацию в обратном порядке (с РНК на ДНК), что и позволяет использовать pegRNA в качестве шаблона для генетических правок.
В целом все это работает в клетке следующим образом. Когда нуклеаза Cas9n садится на геномную ДНК, она расщепляет одну из ее цепочек; после этого в образовавшуюся брешь нужно вставить новый ген, синтезирующийся обратной транскриптазой по матрице шаблона из pegRNA. Именно так сама редакция и происходит: на выходе мы имеем уже одну обновленную ДНК-цепь с исправленным генетическим дефектом. Далее наступает очередь второй ДНК-цепочки, и здесь есть разные варианты.
Дело в том, что существует множество модификаций PE, каждая из которых обладает разными свойствами. Что касается PE, PE2 и PE4 — они разрезают и затем модифицируют только одну цепь ДНК, вторую же «берут на себя» внутриклеточные системы репарации, призванные в данном случае устранить возникшую «асимметрию». Для этого они могут либо вырезать неотредактированную цепь, достроив на ее месте новую по принципу комплементарности (в этом случае редактирование генома будет успешным); либо же первая измененная цепь будет удалена (здесь уже редактирование не выполнено).
То есть, как видим, есть существенный минус и у PE — он попросту срабатывает не всегда… Впрочем, PE4 (как, кстати, и PE5, см. ниже) содержит особые белки, способные ингибировать ошибочную репарацию (mismatch repair inhibiting proteins), что снижает вероятность подобных событий.
Если же речь о PE3 и PE5, то они надрезают и вторую нить ДНК, что улучшает ее репарацию и в целом повышает эффективность редактирования. Однако варианты, разрезающие обе нити ДНК, страдают теми же «болезнями», что и традиционный CRISPR: из-за формирования двойных разрывов могут спровоцировать нежелательные индели. Существуют также и некоторые другие экспериментальные разновидности «праймовых» редакторов: PEMAX — с увеличенной скоростью редактирования; PE6 — со значительно повышенной эффективностью, twinPE — со способностью интегрировать в геном более крупные гены.
В целом, большинство экспериментальных PE имеют как свои достоинства, так и недостатки, а что из них лучше — покажет только время.
«Мостиковое» редактирование: Bridge editing (BrE)
Следующая экспериментальная технология, которую хотелось бы рассмотреть — Bridge editing: пока что она на начальных этапах изучения, но сама по себе сулит завораживающие возможности в будущем, представляя совершенно уникальный инструмент с принципиально инновационным механизмом действия.
Новая технология редактирования использует естественную способность мобильных генетических элементов, известных также как «прыгающие гены» (jumping genes), «вырезать и вставлять себя» по всему геному . Один из них — IS110 — обладает совершенно особыми свойствами: «выпрыгивая» из генома, он образует кольцевую структуру; причем, создавая такую «петлю», этот элемент формирует и промотор — последовательность, с которой может начаться белковый синтез.
Про «прыгучие гены» на «Биомолекуле» также написано уже немало: кому интересны подробности, приглашаем ознакомиться с соответствующим разделом.
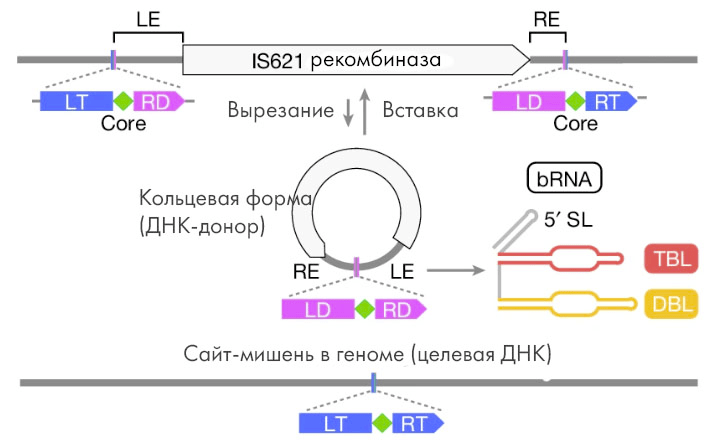
Продуктом такого синтеза в данном случае является «мостиковая» РНК (bridge RNA; bRNA) — она некодирующая, так что дальше транскрипции белковый синтез не идет. bRNA очень похожа на «обычную» направляющую (gRNA), но может распознать не одну цепь ДНК, а сразу две: целевой участок для редактирования и новый ген из донорской ДНК. Затем она привлекает фермент рекомбиназу, который и осуществляет фактическое редактирование. Он связывается с bRNA и катализирует рекомбинацию между донорской ДНК (содержащей целевой ген) и геномным сайтом вставки, что и приводит к желаемому результату: требуемая последовательность интегрируется в нужное место.
bRNA можно запрограммировать на вставку, «вырезку» или инверсию самых разных геномных последовательностей, а потенциал тут выше, чем у «обычных» CRISPR: можно вставить последовательность длиной до 5000 пар оснований .
Это примерно соответствует заявленному для другой вышеупомянутой экспериментальной технологии — twinPE (два этих метода немного схожи). «Двойное» прайм-редактирование использует сразу две направляющие pegRNA в сочетании с сайт-специфической сериновой рекомбиназой, что позволяет, не делая двойных разрывов, интегрировать в геном материал размером >5000 пар оснований, а также инвертировать последовательности величиной до 40 000 пар оснований; удалять же из генома последовательности ДНК он может в диапазоне от 50 до >100 000 пар оснований. Для сравнения: «обычный» PE может делать точечные вставки размером лишь до 40 пар оснований и делеции не более 80 пар.
При этом у BrE отсутствуют риски ошибочного редактирования из-за образования двойных разрывов — их она попросту не формирует. Да и весь механизм здесь проще и точнее: редактирование осуществляется самой BrE всего в один шаг, из-за чего не требуется подключение репарационных систем клетки (и нет связанных с этим рисков). Конечно пока что эта технология все еще очень «сырая» (апробирована лишь на бактериях), но в случае «докрутки» может занять важное место в ряду редактирующих инструментов (и даже «подвинуть с пьедестала» многие из них).
РНК-редактирование: RNA-editing (RE)
заботится о том, чтобы ненароком не дестабилизировать геном, предлагая направить всю мощь редактирующих инструментов на изменения не ДНК, а матричной РНК. Терапевтический эффект тут связан с изменением в «конечном продукте» — то есть, в белковой молекуле. А этого можно достичь, внося изменения как в ДНК, так и в синтезируемую на ее матрице РНК. В последнем случае геном останется нетронутым, а потому если с редактированием что-то пойдет не так, последствия должны быть менее плачевными.
Так-то оно так, только есть недостаток и у такого подхода. Дело в том, что мРНК — продукт промежуточный и временный, короткоживущий. Поэтому здесь для достижения стойкого терапевтического эффекта требуется непрерывно экспрессировать в клетке всю редактирующую РНК машинерию, а это само по себе несет риски нецелевых эффектов. Либо придется вводить продукт многократно, что снижает его привлекательность по сравнению с подходами на основе редактирования ДНК, которые обещают излечение одним уколом.
Несмотря на это, RE-таки сумели «приспособить» к редактированию клеток человека, и сейчас уже эту технологию даже используют в клинических исследованиях (причем подход здесь инновационный и крайне интересный, см. ниже).
Редактирование эпигенома: Epigenome editing —
это в каком-то смысле логическое продолжение предыдущего подхода: попытка изменить экспрессию генов, не затрагивая при этом первичную последовательность ДНК (дабы не дестабилизировать геном). Для этого вносятся мутации в оба расщепляющих нуклеазных домена Cas9, с целью лишить ее целевой ферментативной активности.
В итоге получаем такую «мертвую» нуклеазу (death Cas9 — dCas9): этот фермент уже не «режет», но по-прежнему обладает уникальной способностью «садиться» на ДНК (что и используют ученые). С dCas9 можно соединять различные эффекторные белки, вызывающие эпигенетические модификации и контролирующие экспрессию целевых генов. Это включает в себя такие классические для эпигенетики изменения, как модификации гистонов и метилирование ДНК, что потенциально может помочь в лечении тех заболеваний, на которые такие факторы влияют.
Редактирование эпигенома не сразу «вышло из тени» доклинических исследований, хотя мощный потенциал этого вида терапии был понятен уже довольно давно. Сейчас уже этот инструмент начали изучать и в клинических испытаниях, но поскольку мы лишь в начале этого пути, перспективы его одобрения для использования у людей пока ясны не до конца.
Мы здесь немного за скобками оставляем выбор способа доставки инструментов генного редактирования в клетку, ведь этот вопрос уже разобран в предыдущей статье. Скажем лишь, что в клинических исследованиях долгое время доминировали способы доставки с помощью различных вирусных векторов (см.: «Время первых: как аденоассоциированные вирусы стали лучшими в доставке генов in vivo» и «Лентивирусные векторы: как они стали лучшими векторами для терапии ex vivo»), но в последнее время на пятки им вовсю наступают невирусные методы (такие, например, как липидные наночастицы, хорошо показавшие себя во время ковидной пандемии, где они использовались в составе вакцин Pfizer/BioNTech и Moderna для доставки мРНК спайк-белка), которые, обладая рядом преимуществ, могут со временем найти более широкое применение.
Больше о генном редактировании и его использовании в современной медицине вы узнаете из нашей новой статьи!