А вот ещё один аргумент, на этот раз поближе к генетике:
Fornarino S., Pala M., Battaglia V., Maranta R., Achilli A., Modiano G., Torroni A., Semino O. and Santachiara-Benerecetti S.A. Mitochondrial and Y-chromosome diversity of the Tharus (Nepal): a reservoir of genetic variation // BMC Evolutionary Biology. — 2009. — 9:154:
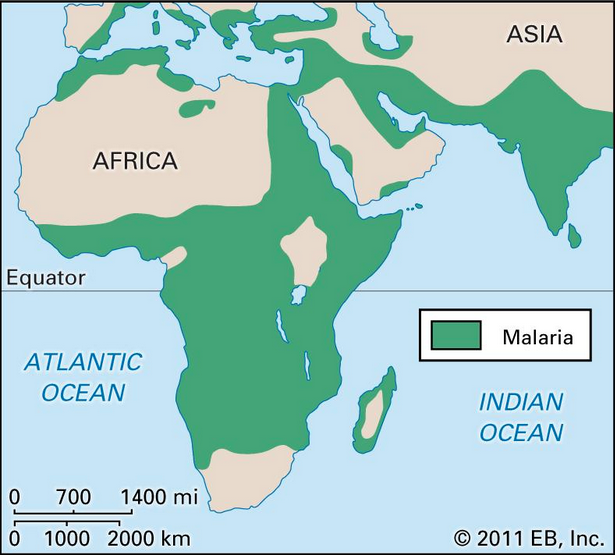
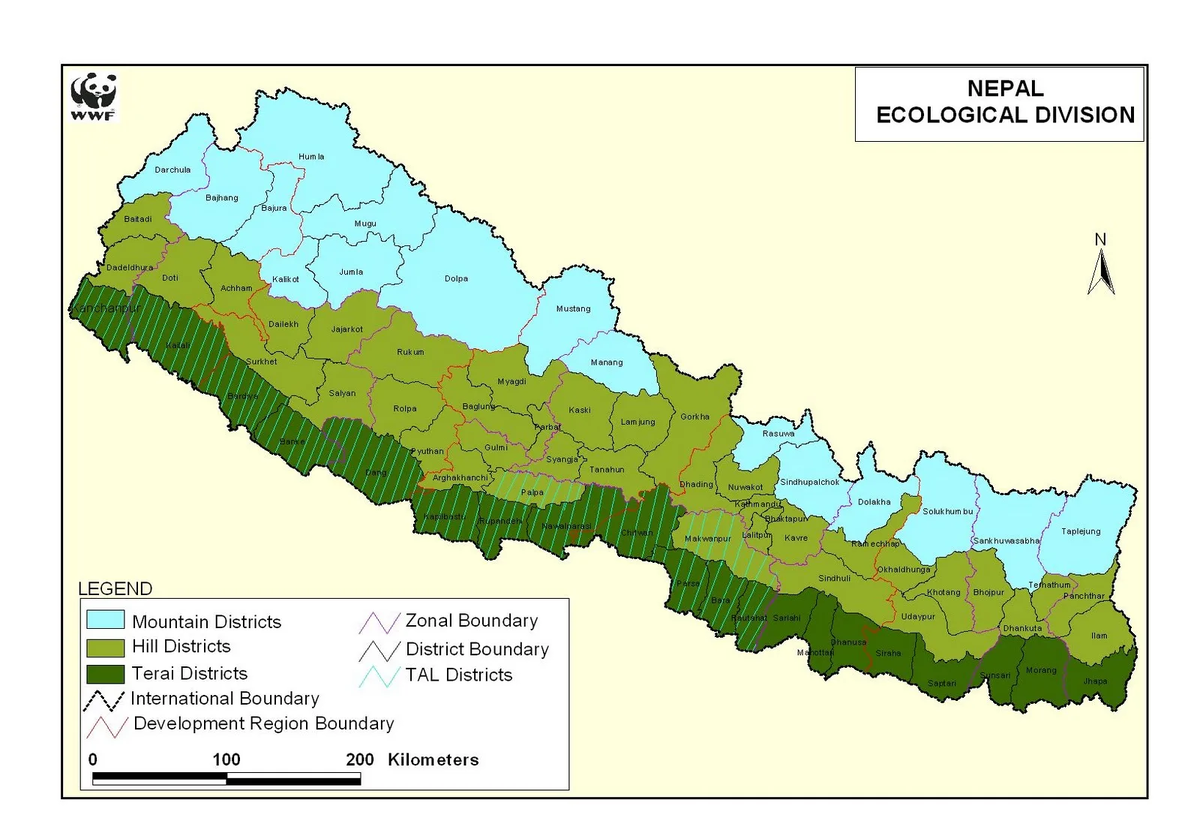
«Тераи, высоко малярийный регион Южного Непала, граничащий с Индией (Рисунок 1), до последних десятилетий, когда малярия была уничтожена, был населён почти исключительно Тхару (Tharu), одним из древнейших и самых многочисленных туземных народов Тераи.
Эта группа известна своею сопротивляемостью малярии, что подтверждается меньшей заболеваемостью малярией в сравнении с проживающими здесь же (sympatric) непальскими популяциями [1], — феномен, не полностью выясненный на генетическом уровне.
Только после по существу полного уничтожения малярии с помощью программы контроля над малярией, запущенной в 1956, несколько других непальских популяций мигрировали и обосновались в Тераи.
Тхару проживают на всём протяжении [территории] страны (в основном в северной полосе Тераи) в деревнях совсем рядом с или даже внутри ранее малярийных лесных зон.
Несмотря на очень глубокую культурную и лингвистическую разнородность, они считают себя уникальным племенным образованием (entity), подразделяющимся на три основные группы (западную, центральную и восточную)…
Образцы состояли из 173 ДНК Тхару из заборов крови мужчин, собранных более 25 лет тому назад, вскоре после значительной миграции других популяций в Тераи вслед за уничтожением малярии, и 104 индийцев. Образцы Тхару состояли из трёх групп из разных деревень: двух в округе Читван в Центральном Тераи (Th-CI и Th-CII) и одной в округе Моранг в Восточном Тераи (Th-E) (Рисунок 1). Индийские образцы также состояли из трёх групп: индусов из Тераи (H-Te, собранных в округе Читван), индусов из Нью Дели (H-ND) и племенные из Андхра Прадеш (T-AP). Отсутствие близкого родства между людьми обеспечивалось данными собеседования…
На основании известного или предполагаемого происхождения гаплогрупп для Y-хромосомы… могут быть установлены три главных компонента (Восточно-Азиатский, Западно-Евразийский и Индийский)… Компонент Индийского субконтинента включает в себя линии гаплогрупп C, F, H, L, O, R и среди индийцев от варьируется от 80% в образцах Нью Дели до 85% в Тераи и до 90% в Андхра Прадеш…
Гаплогруппы H и R являются наиболее часто встречающимися гаплогруппами этого компонента. Гаплогруппа H (Тхару: 25.7%; индийцы: 18.3%) представлена пятью подгруппами: H-M69*, H1-M52*, H1-M370*, H1-M82* и H2-APT. В то время как H-M69* была зафиксирована в схожих пропорциях (в среднем 8.8%) во всех подгруппах Тхару и в двух индийцах Андхра Прадеш (6.9%), H1-M82* наблюдалась во всех Тхару и индийцах. По контрасту, H1-M52* (2.0%) и H1-M370* (6.1%) налюдалась только у индусов Нью Дели, а H2-APT (11.7%) — только у Тхару-CII.
Гаплогруппа R, помимо единичной R* из Нью Дели, была зафиксирована во всех группах как R1a1-M17* и R2-M124 с важными различиями между Тхару (13.5%) и индийцами (52.9%), в основном из-за R1-M17* (8.8% против 41.3%)…»
«Гаплогруппы… R1-M17 и R2-M124… демонстрируют в пределах Индии значительную частоту и гаплотипы, ассоциируемые с высоким микросателлитным разнообразием…
Высокая частота гаплогруппы R1-M17, обнаруженной на территории Центральной Евразии (Central Eurasian territory), наряду с её градиентом распространения, который связывался с индоевропейской экспансией [74-76], должна была бы сохранить некоторую неуверенность по поводу её географического происхождения.
Однако высокое микросателлитное разнообразие подкрепляет древность присутствия, датируемого в наших образцах более 14 тысячами лет [см. Дополнительный файл 3], маркёра M17 на Индийском субконтиненте, как предполагалось Kivisild et al. [11], и подтверждалось Sengupta et al. [15] и Thanseem et al. [71]…
Таким образом, разумно полагать, что даже эта межрегиональная гаплогруппа имеет древние связи с территорией Индии. Интересно, что Y-хромосомы M17 Индийского субконтинента отличаются от таковых в Центральной Евразии (Central Eurasia) тем, что они буквально все — 49a,f/TaqI Ht 11 [77].»
Я осведомлён о проблемности ДНК-датировок. Но вот что меня как неспециалиста в области генетики заинтересовало.
Откуда у туземцев в малярийном регионе[1] на южных склонах Гималаев, куда по определению до самого недавнего времени не могли проникнуть не то что неиндийцы, но даже жители самих равнин Южной Азии, такой набор мужских гаплогрупп семейства R, особенно R1a1-M17*?
И второй момент, мимо которого я не смог пройти: название статьи — Tharus (Nepal): a reservoir of genetic variation. Резервуар генетического разнообразия.
Все южные склоны Гималаев именно в силу своей изрезанности, фрагментации на относительно изолированные горные долины, с одной стороны, и в силу вертикальной поясности своих природно-климатических зон, — с другой, несомненно, представляют собою резервуар не только генетического и биологического (мы тут находим, например, медведя, волка, гималайский кедр, берёзу и т.д. и т.п.), но и этнолингвистического и культурного разнообразия.
Ещё Л.Д. Калла отмечал, что именно это способствовало формированию родственных, но в то же время отличных друг от друга ИЕ [индоевропейских] диалектов. И относительное перенаселение заставляло их носителей спускаться вниз на равнины и тем самым служило естественным механизмом миграции.
[1] Малярийный аргумент совершенно игнорируется сторонниками европейского или степного происхождения индоиранцев, хотя он ещё раз указывает на категорическую (на этот раз эпидемико-биологическую) невозможность выведения ариев из не поражённых малярией регионов Евразии.
Между тем «вполне вероятно предположить, что
индо-арийские племена, так же как греческие и италийские, пришедшие соответственно, в южные области европейского континента, должны были обладать иммунитетом к малярии и её летальных последствий [так в тексте. — А.С.]. Такой иммунный механизм мог быть приобретён этими племенами только в областях исторического распространения тропической малярии,
в частности, в Древней Малой Азии и других областях Восточного Средиземноморья, где был распространён
генетически передаваемый вид анемии — талассемия (средиземноморская болезнь), ген которой в гетерозиготном состоянии увеличивает сопротивляемость организма к малярии (при летальности этого заболевания с геном талассемии в гомозиготной аллели).
Если предположить, что популяции индо-арийских племён, вторгшихся в Индостан, не имели этих генетически выработанных способов резистентности к малярии, они не смогли бы выжить в таких условиях.
Поэтому переселение этих популяций в Индостан с запада (из переднеазиатского ареала талассемии) представляется более вероятным, чем традиционно предполагаемая миграция с севера из Центральной Азии, где нет этого вида генетически передаваемой анемии».
(Гамкрелидзе Т.В. Новая парадигма в индоевропейском сравнительно-историческом языкознании: к реконструкции индоевропейского праязыка и индоевропейской прото-культуры // Труды Отделения историко-филологических наук РАН / отв. ред. А.П. Деревянко. — М.: Наука, 2005. — С. 19–20.)
Однако столь же возможна и миграция индоевропейских народов с востока из ирано-индийского пограничья на запад в Переднюю Азию.
Вот что говорится в Википедии (https://ru.wikipedia.org/wiki/Талассемия):
"В зависимости от того, синтез какого из мономеров нарушен, разделяют альфа-, бета- и дельта-талассемию. По тяжести клинических проявлений выделяют тяжёлую, среднюю и лёгкую формы заболевания. Альфа-талассемия. Связана с мутациями в генах HBA1 и HBA2. Есть всего 4 локуса, кодирующего α-цепи. Наличие мутации в одном из локусов приводит к минимальным клиническим проявлениям. Нарушения в двух локусах выражаются лёгкой формой анемии. При мутациях в трёх локусах возникает значительное уменьшение продукции α-глобина. При этом избыточные цепи β-глобина образуют тетрамеры — гемоглобин Н. Эта форма носит также название гемоглобинопатии Н. Характер заболевания может варьироваться от лёгкой до тяжёлой картины гипохромной микроцитарной анемии. Присутствие мутаций во всех четырёх аллелях альфа-глобина несовместимо с жизнью. Ребёнок с такой патологией погибает внутриутробно или вскоре после рождения. Из пуповинной крови таких детей можно выделить гемоглобин Барта. Этиология. Талассемию вызывают точечные мутации или делеции в генах гемоглобина, ведущие к нарушению синтеза РНК, что приводит к уменьшению или полному прекращению синтеза одного из видов полипептидных цепей. Синтез цепей другого вида продолжается. Это приводит к образованию нестабильных полипептидных агрегатов из избыточных цепей, нарушающих нормальное функционирование эритроцитов и их разрушению. Повышенный гемолиз эритроцитов вызывает анемию. Эпидемиология.
Альфа-талассемия распространена в Западной Африке и Южной Азии.
Бета-талассемия часто встречается в странах Средиземноморья, Западной Азии и Северной Африки.
Это регионы, где распространена малярия. Гетерозиготные носители мутаций в генах альфа- и бета цепей гемоглобина являются более устойчивыми к малярийному плазмодию.
Имеются очаги талассемии в Азербайджане, в равнинных районах которого гетерозиготная бета-талассемия наблюдается у 7–10 % населения."
Даже автохтоны Южной Азии становились жертвами малярии:
«Одной причиной упадка городской жизни в Мохенджо-даро было, вероятно, слабое здоровье его жителей. Скелеты из верхних слоёв, оставленные в заброшенных домах и на [покинутых] улицах… продемонстрировали следы того, что многие люди страдали от малярии, распространявшейся комарами, вероятно, размножавшимися на затопленной наводнениями земле неподалёку от города» (McIntosh J. The ancient Indus valley: new perspectives. — Santa Barbara: ABC–CLIO, 2008. — P. 396.).
Насколько же надо игнорировать медико-эпидемиологические факты, чтобы продолжать настаивать на выведении предков индоиранцев из регионов, не заражённых малярией?!
Материал издан в публикации дискуссии о прародине индоевропейцев на Генофонд.рф весной 2015 г.: AIT versus OIT: дискуссия на "Генофонд.рф" (составление, корректура, редактирование, предисловие, сноски и приложения Семененко А.А.) / А.А. Семененко, И.А. Тоноян-Беляев, Е.В. Балановская, О.П. Балановский, Я.В. Васильков, В.В. Запорожченко, Л.С. Клейн, А.Г. Козинцев, С.В. Кулланда, В.А. Новоженов. — Воронеж: На правах рукописи, 2015. — 846 с. — ISBN 978-5-9907439-0-8. — С. 775—772.
Автор публикации — дипломированный историк со специализацией по кафедре археологии и истории Древнего Мира и степенью кандидата исторических наук.



















