Первая часть этой темы по этой ссылке.
Для удобства опять приведу матрицу, которая служит примером для данного случая.
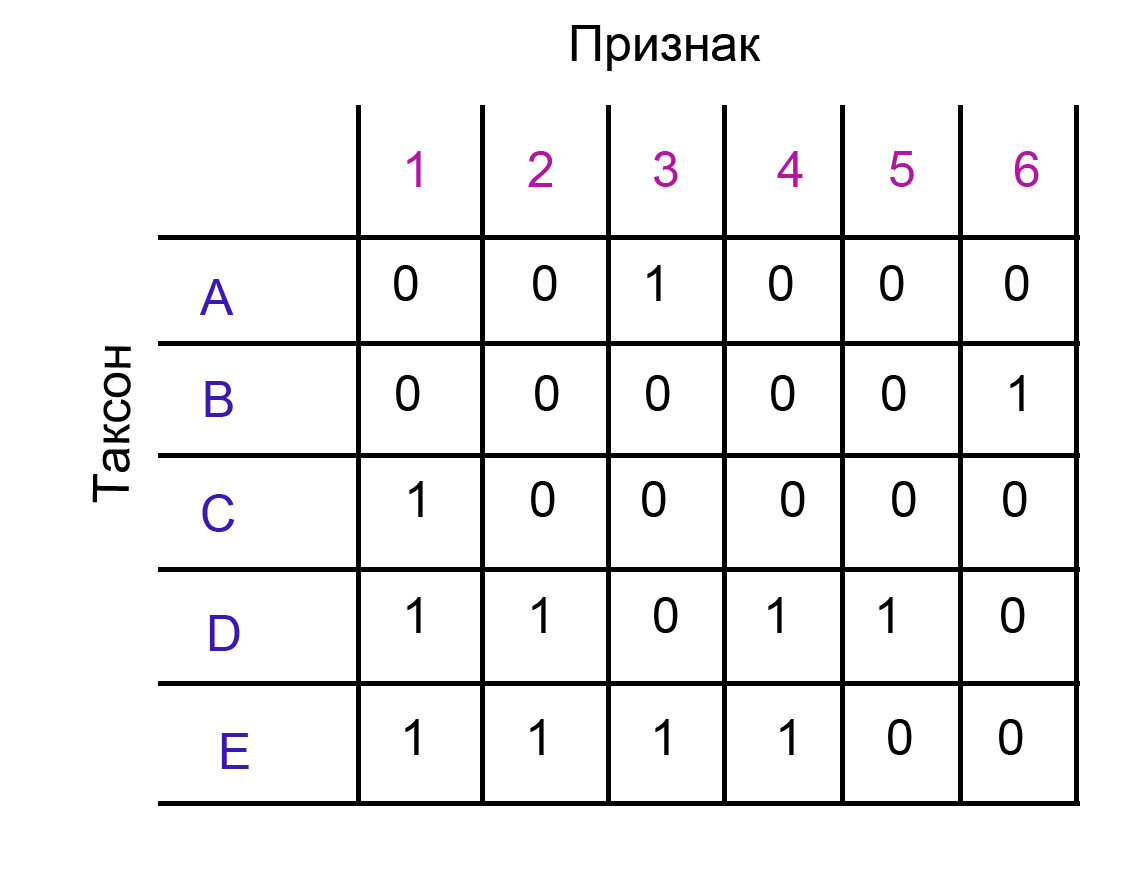
Я отвечу на два вопроса, которые, скорее всего, появятся у некоторых читателей.
1) Почему после того, как мы развесили первый признак на филогению, нельзя просто выбрать само короткое (экономное) дерево, а все остальные варианты отбросить? И так каждый шаг, пока мы не развесим все признаки?
Этот вопрос вполне справедлив, потому что в прошлом посте после добавления каждого признака я выбирала самое короткое дерево. Таким образом, у меня получилось прийти к самому короткому дереву после добавления всех признаков. Вот оно. У него 7 шагов, что означает, что его длина - 7.
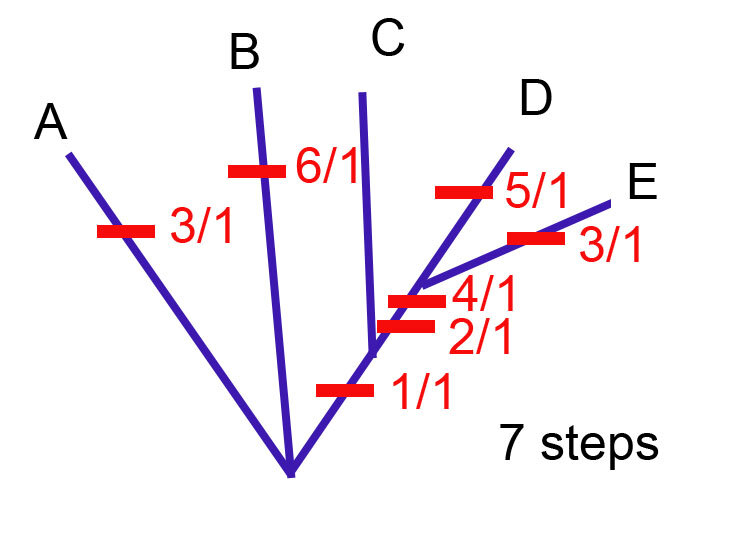
И если вы каким угодно способом обсчитаете эту матрицу методом максимальной парсимонии, у вас получится именно такое самое экономное дерево. Все остальные варианты будут длиннее.
К сожалению, в преобладающем большинстве случаев так ничего не получится. В данном случае, я просто начала с того признака, который позволил нам в итоге прийти к самому короткому дереву. Учитывая, что признаков было мало, и самое короткое дерево всего одно, это было вполне реально. Однако в реальности матрицы обычно гораздо больше, и одинаково коротких деревьев обычно более одного, и чтобы их всех найти, надо ходить разными путями. Но даже если самое короткое дерево всего одно, его бывает не так просто найти.
Даже в нашем простом случае это можно легко продемонстрировать. Например, начав с признака 3, который объединяет таксоны A и E. Вот это самое короткое дерево с одним шагом.
Теперь давайте постепенно добавлять все остальные признаки в порядке 1, 2, 4, 5, 6 и выбирать каждый раз самое короткое дерево.
В итоге окончательное дерево имеет длину 9, что на 2 больше, чем, когда мы начинали с признака 1.
Поскольку при работе с более или менее осмысленными матрицами, мы не будем знать, с какого признака начать, то придется перебирать все деревья.
2) И все же, почему же признаки, в которых только одно состояние присутствует у более, чем одного таксона, ничего не группируют? Например, признак 6 вполне может группировать таксоны A, B, C, E относительно таксона D. Более того, наше самое короткое дерево, то самое, что я привела в начале, вроде как не полностью разрешено. И если признак 6 поместить немного иначе, то можно добиться полного разрешения, поскольку он как раз и будет группировать таксоны A, B, C, E относительно таксона D. При этом длина дерева не изменится.
Приведу два альтернативных варианта.
Для того, чтобы понять, что это не имеет никакого смысла, и даже создает иллюзию, что у нас есть как бы клада A, C, D , E, про которую на самом деле нам ничего не может сказать приведенная матрица, обратимся опять к теме укоренение. До этого я обсуждала его только применительно к топологии. Давайте теперь посмотрим, что происходит при этом с признаками при укоренении.
В прошлом посте я просила вас помнить, что мы работаем с неукорененным деревом, и укореняем его в конце. В данном случае корнем может быть любой таксон.
Даже если мы укореним на таксон B, то при укоренении, он автоматически будет противопоставляться всем остальным таксонам. Так что признак 6 нам ничего не скажет нового об отношениях между таксонами. Таким образом, есть два варианта насчет группы A, C, D , E: (1) мы ее задали до анализа, заранее решив, что будем укоренять на таксон B. В этом случае она просто получится автоматически. Таким образом, вводить признаки специально для нее нет смысла. (2) Мы укореняем на другой таксон (внутри нее), и эта группа неизбежно развалится и никаких апоморфий для нее быть не может.
Давайте посмотрим, что будет, если мы укореним, к примеру, на таксон D.
Как видите, состояние 6/1 - ничего более, чем апоморфия для одного единственного таксона B.
Обратите внимание, как изменилась топология и распределение признаков по сравнению с предыдущей картинкой, хотя это всего одно дерево, только по-разному укорененное. При укоренении надо будет заново развесить признаки, убедившись, что это сделано самым экономным способом. Длина дерева при этом поменяться не может.
Так что, действительно, если дерево не укоренено, то более корректно показывать в основании политомию (как это сделано в первом дереве в этом посте), чем выбирать апоморфию для корня и делать вид, что это апоморфия для всех таксонов, кроме корня. Потому что даже если в природе эта гипотеза совпадает с реальностью, протестировать это можно будет, только если будет хотя бы еще один таксон, у которого будет такое же состояние, как и у корня.