В связи с проведением строительных работ в провинции Хэйлунцзянь Китая были обнаружены останки древних людей, что позволило ввести в оборот данные еще по 25 объектам возрастом от 33,4 до 3,6 тысяч лет (тл). Исследования китайских ученых выявили интересную аналогию развития человеческих общностей в бассейне Амура и в Европе.
Место находок и новые образцы
Провинция Хэйлунцзянь расположена на северо-востоке Китая, граничит с территорий России, отделенная от нее рекой Амур.
Основным объектом раскопок стала равнина между рекой Сунгари, впадающей в Амур, и ее притоком Нэньцзян (Нен).
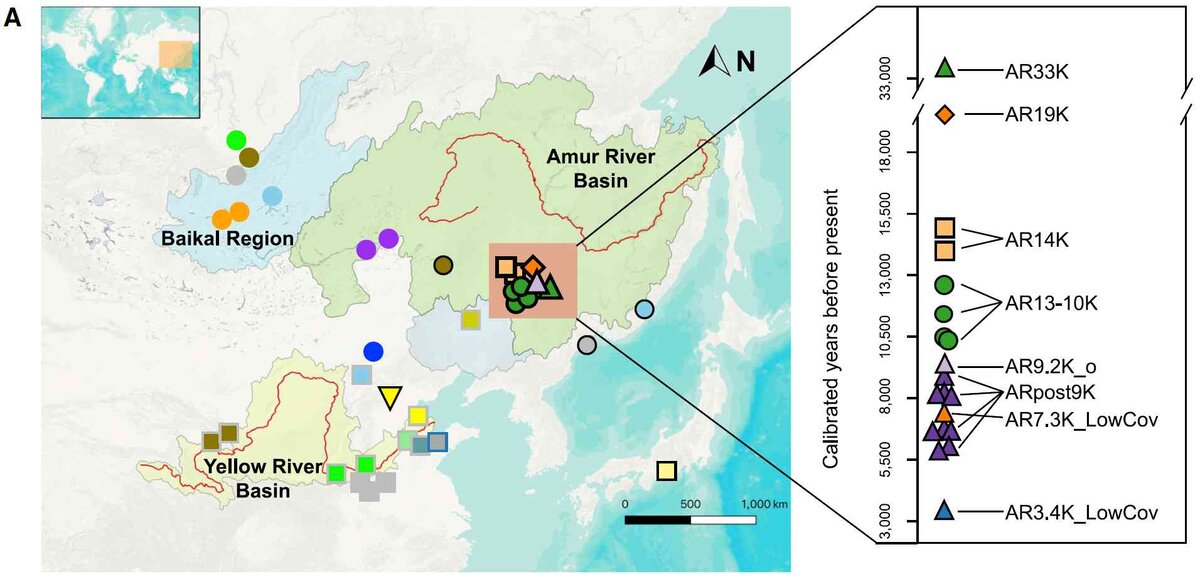
Среди новых находок останки женщины (AR33K)возрастом от 34?3 до 32,4 тл, это четвертый образец старше 30 тл в северной части Восточной и Северо-Восточной Азии. Большой интерес представляют останки мужчины (AR19K, 19,6-19,2 тл, Y-группы С2 и мтДНК группы G2), жившего в конце последнего ледникового максимума (ПЛМ). Интересны и другие образцы, так как они восполняют пробелы в истории людей в Приамурье и в целом в Восточной Азии.
В целом образцы группируются следующим образом.
Соотношение между самыми древними образцами
Несмотря на существенную разницу в возрасте амурский AR33K демонстрирует сходство с человеком из пещеры Тяньюань (возраст 40 тл), эти образцы формируют отдельный от других кластер. Меньшее сходство у этого образца с образцами примерно одинакового с ним возраста, с образцом с Янской стоянки и из Сальхута в Монголии, хотя последнего рассматривают как генетическое смешение компонент из Тяньюань (75%) и с Янской стоянки (25%).
Интересно отметить, что, в отличии от образца из Тяньюань, AR33K не демонстрирует схожести с европейским GoyetQ1 16-1 (35 тл из Бельгии).
Авторы делают вывод, о том, что 40-33 тлн генетический компонент Тяньюань/AR33K был широко распространен на севере Восточной Азии. Хотя потомков среди современных людей не имеет.
Поэтому интересен образец AR19K как некое переходное звено. Анализ этого образца важен для изучения популяционной динамики северного региона Восточной Азии. При оценке методом главных компонент и f3-статистики была выявлена его большая близость с более поздними популяциями Восточной Азии. Это показывает, что начало популяционных изменений на севере Восточной Азии относиться ко времени не позже конца ПЛМ. Таким образом, генетическая дифференциация между югом и севером Восточной Азии началась на 10 тл раньше, чем предполагалось до этого.
Исследование достаточно большое количество образцов 15-10 тл позволило авторам сделать ряд выводов. Формирование генетически однородной популяции началось не позже ПЛМ. Представителем этой новой популяции является уже AR19K. Причем эта популяция отличается от более южных популяций, таким образом, во время ПЛМ, а возможно и раньше, уже начался процесс разделения северных т южных популяций Восточной Азии.
Начиная с 14 тлн, отмечается увеличение эффективной численности популяции, авторы связывают это с переходом к производящему хозяйству, в частности с выращиванием просо.
Амур – Сибирь – Европа
Изменения происходящие в амурской популяции, существовавшая 40-33 тлн популяция не оставила прямого потомства. Во время или перед ПЛМ происходит замена ее на другую генетическую общность, а затем около 14 тлн рост численности популяции. Авторы отмечают, аналогичную ситуацию в Сибири, появление новой мощной популяции потомков Y-гаплогруппы R, причем тоже во время или до ПЛМ. А после ПЛМ рост численности активная миграция популяций потомков.
Аналогичная ситуация в Европе. Первоначальная популяция до 40-39 тлн, не оставившая потомства, затем происходит формирование новой генетической общности. Во время или до ПЛМ появляется новая Y-гаплогруппы I и после ПЛМ происходит широкое распространение ее субкладов.
Можно предположить, что аналогия обусловлена следующим. Миграцией в новую экологическую среду полиморфной популяции. Под воздействием новых условий среды происходит смена доминантных групп в популяции. Вероятность нахождения останков представителя старой доминантной группы выше, просто из-за их большей численности. Постепенно картина меняется, кроме того, неизменные условия среды ведут росу генетической однородности общности. Приближение ПЛМ ускоряет такие процессы, а более благоприятные условия существования после него ведут к росту численности общности.
Статья The deep population history of northern East Asia from the Late Pleistocene to the Holocene, первый автор Xiaowei Mao