Скелетные мышечные волокна являют собой многоядерные клетки, которые содержат в основном миофибриллы, окруженные водной средой, именуемой саркоплазма. Некоторые данные свидетельствуют о том, что саркоплазматическая гипертрофия, или непропорциональное расширение саркоплазмы по отношению к миофибрильной аккреции белка, совпадает с ростом мышечных волокон или тканей во время применения силовой тренировки. Существуют также данные, подтверждающие другие формы гипертрофии, возникающие во время периодов силовой тренировки, включая пропорциональное увеличение миофибриллярных белков с ростом волокон или тканей (т. е. обычная гипертрофия), или рост белка миофибрилл, предшествующий росту волокон или тканей (т. е. упаковка миофибриллы). В этом обзоре будут обсуждаться методы, которые были использованы для исследования этих режимов гипертрофии, в то время, как Особое внимание уделяется саркоплазматической гипертрофии. Таким образом, будут положены описания, этого процесса, а также широкие последствия этого явления.
Вступление
Исследования, начиная со второй половины 20-го века и до наших дней, показали, что силовая тренировка увеличивает поперечный рост скелетных мышечных волокон (обзор в Folland and Williams, 2007; Walker et al., 2011; Haun et al., 2019c). Однако меньше известно о морфологических адаптациях, которые происходят в мышечных волокнах после проведения силовых тренировок. Группа ученых недавно предположила, что силовая тренировка, выполняемая с большим объемом приводит к большему росту волокон за счет непропорционального увеличения объема саркоплазмы по сравнению с аккрецией белка миофибрилл (Haun et al., 2019b; Vann et al., 2020b).
Саркоплазматическая гипертрофия была основным описанием этого явления, и этот термин будет использоваться далее. Эти исследователи далеко не первые, кто предположил наличие саркоплазматической гипертрофии, возникающей в ответ на силовую тренировку , и исследования, приведенные ниже в этом обзоре, отчасти пытались подтвердить этот механизм. В научном сообществе принято считать, что обычная гипертрофия возникает в ответ на силовую тренировку ; в частности, мышечные волокна подвергаются росту своего поперечного сечения, вследствие силовой тренировки, за счет пропорциональной аккреции миофибриллярного белка и пространства саркоплазмы. Это означает, что, если человек демонстрирует увеличение средней площади поперечного сечения волокон (fCSA) на 20% после выполнения программы силовых тренировок, то так как миофибриллы составляют ∼85% внутриклеточного пространства, 17% этого пространства будут занимать миофибриллы ,тогда как на саркоплазму приходится 3% объема волокон.
Этот способ измерения гипертрофии мышечных волокон, безусловно, имеет смысл. Несмотря на это, на сегодняшний день существует только одно исследование на людях, которое слабо поддерживает этот механизм (Luthi et al., 1986), и это исследование будет обсуждаться далее более детально.
Многочисленные исследования показали, что скорость синтеза миофибриллярного белка увеличивается через несколько часов после выполнения силовой тренировки (обзор Walker et al., 2011; Haun et al., 2019c ,обзор в Damas et al., 2015). Эти данные подтверждают традиционную модель гипертрофии, рассмотренную выше. Несмотря на свою информативность, некоторые исследования были более противоречивыми. На самом деле, увеличение скорости синтеза миофибриллярного белка после тренировки не всегда бывает связано с долгосрочными гипертрофическими исходами (обзор Mitchell et al., 2015) без учета других переменных.Поэтому исследования, изучающие интегрированные скорости синтеза миофибриллярного белка в течение нескольких дней или недель от начала тренировки, дали лучшие ассоциации между повышением миофибриллярного синтеза белка и конечными гипертрофическими исходами (Brook et al., 2015; Damas et al., 2016a; Bell et al., 2019).
Цель данного обзора состоит в том, чтобы представить подробный и углубленный обзор сценариев, в которых саркоплазматическая гипертрофия наблюдалась и не наблюдалась в литературе в ходе силовых тренировок у человека.
Внутриклеточная морфология мышечных волокон.
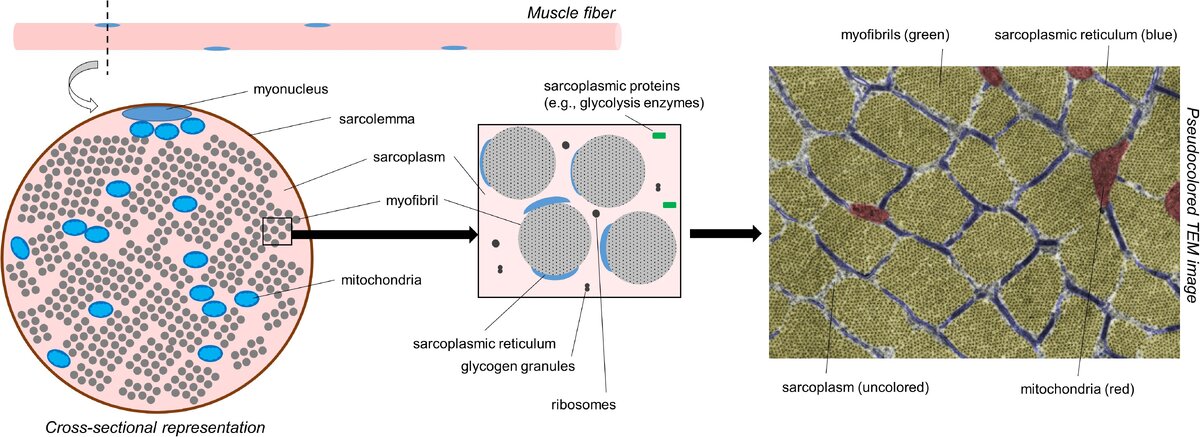
Прежде чем обсуждать различные формы гипертрофии, важно, чтобы читатели оценили внутриклеточную морфологию мышечных волокон. На рис.1
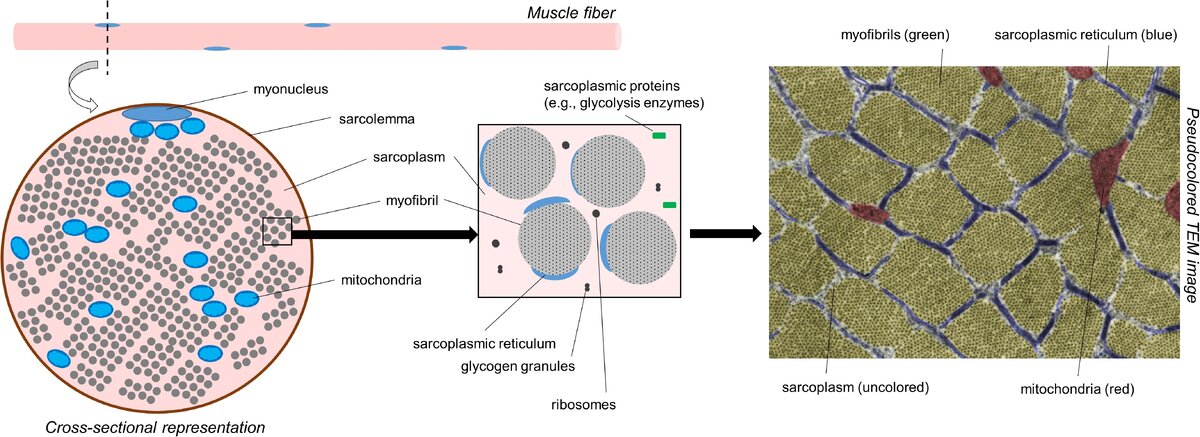
показано поперечное сечение мышечного волокна, как оно выглядит под просвечивающим электронным микроскопом (сокращенно ТЭМ). С молекулярной точки зрения мышечная ткань состоит из 75% воды, 10-15% миофибриллярного белка и 5% не миофибриллярного или саркоплазматического белка (Vann et al., 2020b). С пространственной точки зрения, большая часть мышечного волокна занята миофибриллами; по некоторым оценкам, миофибриллы занимают ∼85% внутриклеточного пространства (Macdougall et al., 1982; Alway et al., 1988; Claassen et al., 1989). Миофибриллы состоят из различных белков, в том числе и миозина тяжелой цепи и легкой цепи изоформ, различных изоформ актина, тропонина и различных изоформ тропомиозина, Z-линия белков, и другие белки, которые служат для поддержания структурной целостности миофибрилл (например, титин и небулин в частности) Haun et al., 2019c; Vann et al., 2020b.
В совокупности этот белковый пул обычно называют миофибриллярной фракцией мышечной ткани. Расстояние между миофибриллами невелико, то есть, по приведенным выше оценкам, ∼15% внутриклеточного пространства не занято миофибриллами. Это пространство содержит органеллы,как и клеточные компоненты, включая митохондрии (в форме ретикулума), саркоплазматический ретикулум, макромолекулы (например, рибосомы, гликоген и липидные капли), а также различные белки и ферменты.
ТЭМ был использован для того, чтобы показать, что митохондрии занимают всего∼5-6% пространства внутри мышечных волокон, в то время как саркоплазма занимает ∼9% (Claassen et al., 1989).
Поэтому водная среда, в которой находятся миофибриллы и не миофибриллярные компоненты, называется саркоплазмой. Сродни цитоплазме в других клетках, саркоплазма имеет решающее значение для поддержания ионного и рН-баланса в мышечных волокнах. Не миофибриллярные компоненты мышечных волокон, а также саркоплазма составляют то, что было названо саркоплазматической фракцией мышечной ткани.
На рис.1
не показано, как различия в типе волокон влияют на морфологию клеток.
Исследования на животных показали, что основные различия между медленными волокнами (тип I) и быстрыми волокнами (тип II) заключаются в том, что волокна типа один содержат больше митохондрий либо в расширении митохондриального ретикулума по сравнению с быстрыми волокнами второго типа (Schiaffino et al., 1970) ,а также в быстро сокращающихся волокнах второго типа имеется больший саркоплазматический ретикулум и везикулы, окружающие миофибриллы (Lee с соавт., 1991) .Кроме того диаметры миофибрилл, по-видимому, несколько больше в волокнах типа два , хотя саркомеры в типе один немного длиннее, а z-линии немного толще (Schiaffino et al., 1970). Некоторые из этих характеристик волокон также были показаны в скелетных мышцах человека с использованием методов электронной микроскопии (ТЭМ) (Alway et al., 1988).
Рисунок 2
- это изображение ТЭМ, которое визуально демонстрирует некоторые из этих различий в типе волокон.
Сарколемма, или клеточная мембрана мышечного волокна, также является важной темой для обсуждения. Сарколемма содержит различные белки, включая мембраносвязанные рецепторы и транспортеры. Сарколемма также содержит трансмембранные белки, и эти белки образуют комплексы, которые связывают внутриклеточные цитоскелетные белки с белками, находящимися во внеклеточном матриксе (Henderson et al., 2017). Последние данные указывают на то, что некоторые из белков сарколеммы- включают в себя фракции миофибриллярных белков (например, дистрофин), в то время как другие-саркоплазматических фракций (например, vinculin) (Vann et al., 2020b). Важно отметить, что хотя эти белки обогащены одной или обеими фракциями, технически они не подпадают ни под одну из этих классификаций. Эта тема будет повторно рассмотрена ниже.
Методы оценки саркоплазматической гипертрофии.
Существует четыре метода, которые использовались учеными для сообщения о том, что саркоплазматическая гипертрофияпотенциально может возникнуть в ответ на силовую тренировку . Три из этих методов включают электронную микроскопию- ТЭМ, окрашивание фаллоидином ,окрашивание SDS-PAGE и Coomassie,и также оценка специфического напряжения в изолированных мышечных волокнах, которая может служить косвенным доказательством саркоплазматической гипертрофии. Механика, преимущества и недостатки каждого метода будут обсуждаться ниже.
1) Метод электронной микроскопии (ТЭМ).
Этот метод включает в себя анализ небольшого кусочка мышечной ткани (в пределах 1-2 мг) . После чего выполняются небольшие срезы, которые помещаются на небольшие сетки, а срезы химически обрабатываются с использованием определенных препаратов (уранилацетата и цитрата свинца) для создания контраста между клеточными структурами. Сетки, содержащие один или несколько разделов, затем можно просмотреть в микроскопе -ТЭМ. Предполагая, что эти методы выполняются правильно, изображения, полученные с помощью этой методики, обеспечивают анализ миофибрилл с хорошим разрешением при увеличении до 1000–20 000 раз. Затем исследователи могут использовать ручное или автоматизированное программное обеспечение для анализа изображений для определения размеров миофибрилл, митохондрий и пространственной "пустоты", существующей между этими структурами (то есть занимаемой саркоплазмой). Миозиновые и актиновые нити также могут быть изучены при увеличении ∼40 000× .
Основным недостатком ТЭМ является то, что методика дает только двумерное изображение части одного или двух мышечных волокон. Хотя несколько изображений могут быть случайным образом получены через несколько мышечных волокон, чтобы получить более целостное представление о ткани, получение 20 изображений на объект (в лучшем случае) даст данные только для частей 20-40 волокон. Второе ограничение ТЭМ заключается в том, что фиксация, обезвоживание, встраивание, окрашивание и.т.д., требуют нескольких дней лабораторной работы. Это не только проблематично с точки зрения пропускной способности, но и большое количество этапов может искусственно изменить расстояние между миофибриллами, что приведет к путанице при измерении саркоплазматического объема (!). В связи с этим были зарегистрированы артефакты при измерении методом ТЭМ (например, расстояние между клеточными структурами) из-за ошибок обработки (Mollenhauer, 1993), и эти явления могут привести к ошибочным выводам о клеточных свойствах (о наличии саркоплазматической гипертрофии). Наконец, устройства ТЭМ редко встречаются в лабораториях физиологии физической культуры, учитывая затраты и опыт, необходимые для эксплуатации и обслуживания данного оборудования.
2) Окрашивание препаратов- Фаллоидином.
Это гистологический метод, при котором окрашивающий раствор, содержащий фаллоидин наносят на мышечные срезы, приклеенные к стеклянным предметным стеклам микроскопа. Химические свойства фаллоидина позволяют ему с высокой специфичностью связываться с актиновыми полимерами; таким образом, одной из сильных сторон метода является специфичность окрашивания. Относительное содержание актина в поперечных срезах мышц, инкубированных с фаллоидином затем может быть получено в целых волокнах ∼20-30 с помощью простой флуоресцентной микроскопии при увеличении в 200×. Если получено несколько изображений разреза в разных областях ткани, то можно получить данные от∼50-100 волокон. С помощью автоматизированного программного обеспечения для подсчета пикселей затем можно обеспечить непредвзятое подсчет зеленых пикселей в пределах границ, окрашенных дистрофином, чтобы получить относительное количество актина на миофибриллу. Помимо высокой специфичности связывания фаллоидина с актином, еще одной сильной стороной этого метода является анализ большего количества целых волокон по сравнению с визуализацией ТЭМ.
Этапы, связанные с обработкой тканей, также менее трудоемки по сравнению с подготовкой ТЭМ. Однако, как и ТЭМ, эта техника также ограничена двумерным изображением. Еще одно ограничение заключается в том, что в мышечных волокнах существуют не сократительные актиновые белки, которые служат каркасными белками (Stromer, 1998). Хотя эти белки, вероятно, не составляют большую часть саркоплазматической белковой фракции, фаллоидин, вероятно, связывается и с этими белками , генерируя сигнал из не контрастного материала. Правильная консервация тканей путем медленного замораживания в среде с оптимальной температуры разреза, также необходима для успешного выполнения этой техники. Неправильное замораживание может привести к часто наблюдаемому артефакту обнаружения больших структур (!), которые могут быть неправильно истолкованы как пространство, не занятое миофибриллами (Meng et al., 2014).
3) Третий метод -это Додецилсульфата натрия-электрофорез в Полиакриламидном геле (SDS-PAGE) с окрашиванием Coomassie.
Это не гистологический метод, который позволяет определить относительное содержание тяжелых цепей миозина , актина, тропомиозина, тропонина и миозина легких цепей белковых изоформ на миллиграмм влажной или сухой мышечной ткани. Учитывая, что выполняются различные итерации этого анализа (Roberts et al., 2018; Haun et al., 2019b; Vann et al., 2020b),далее будет представлено более подробное описание этой техники по сравнению с другими. Также на схеме на рис. 3,https://www.frontiersin.org/files/Articles/542447/fphys-11-00816-HTML/image_m/fphys-11-00816-g003.jpg показывается, как интерпретируются результаты данного анализа. Следует отметить, что другие исследователи также использовали этот метод с целью количественного определения относительного количества сократительного белка на единицу ткани у различных видов животных (Yamada et al., 1997; Thys et al., 2001; Ефремов и соавт., 2008). После получения биопсии мышцы ткань заворачивают в предварительно маркированную фольгу, замораживают в жидком азоте и транспортируют при температуре -80°C для длительного хранения. Учитывая, что ткань заморожена в естественном состоянии, а мышечные волокна содержат ∼75% воды, как было указано ранее, это обычно называют влажной тканью. В течение дня эксперимента ткань удаляют при температуре -80°С и помещают на охлажденную жидким азотом (LN2) ступку. Затем ткань измельчают с помощью охлажденного пестика, и часть ткани (≈20 мг) помещают в предварительно взвешенную пробирку объемом 1,7 мл, содержащую буфер ,так называемый “буфером один”. Эти детали очень важно отметить, поскольку этот первый буфер содержит вещество детергент Triton-X, который способен лизировать клеточные мембраны, не солюбилизируя миофибриллы. Этот буфер также может быть одной из причин того, что некоторые из белков сарколеммы обогащены саркоплазматической белковой фракцией, как указывалось ранее в обзоре. Затем трубки повторно взвешивают, чтобы получить вес влажной мышцы, основанный на разнице веса между трубкой до и после добавления ткани. Трубка, содержащая буфер 1 и ткань, затем удаляется из аналитической шкалы, измельчается с помощью плотно прилегающих пестиков до образования суспензии, после чего трубки закрываются крышками и центрифугируются. После центрифугирования остаются жидкий супернатант и гранулы. Супернатант содержит в основном буфер 1, а также саркоплазматические компоненты (например, саркоплазматические белки, митохондриальные белки). Гранула содержит в основном миофибриллярные белки, учитывая, что они не были солюбилизированны. Однако также обнаруживаются определенные гистоновые белки в гранулированной фракции с помощью протеомики (Vann et al., 2020b). Хотя присутствие этих ядерных белков минимально по сравнению с другими сократительными белками (например, тяжелой цепью миозина, титином, небулином и другими), эти данные предполагают, что ядра, вероятно, гранулируются во время первой стадии центрифугирования. Примерно 90% супернатанта удаляют с помощью пипетки, супернатант помещают в новую пробирку объемом 1,7 мл,и эта пробирка, содержащая саркоплазматическую фракцию, хранится при температуре -80°C для последующего анализа. Затем в исходную пробирку объемом 1,7 мл, содержащую миофибриллярную белковую гранулу, добавляют дополнительный буфер 1, и гранулу повторно суспендируют для промывки гранулы. Полученную суспензию центрифугируют, а полученный супернатант отбрасывают после центрифугирования. Гранулы миофибрилл можно высушить на воздухе на льду, а затем повторно суспендировать в том, что мы назвали буфером 2, который представляет собой буфер на основе Трис, содержащий ингибиторы протеазы, а также хлорид калия, глицерин и спермидин. Примечательно, что все последние добавки способствуют солюбилизации гранул миофибриллы. Как только миофибриллы повторно суспендируются, заданный объем ресуспензии (например, 10 мкл) может быть получен с восстановительным буфером и подвергнут SDS-PAGE. Затем гель может быть окрашен , плотность видных полос, существующих при ∼220 кд (тяжелая цепь миозина) и 43 кд (актин), может быть количественно определена, и плотность полос может быть разделена на мышечную массу, чтобы получить относительные количества тяжелой цепи миозина и актина. В связи с ограничениями, обсуждаемыми с Тэм и гистологией, использование этого метода имеет свои преимущества. Во-первых, отбирается большее количество мышечной ткани по сравнению с окрашиванием ТЭМ или фаллоидином, и это преимущество обеспечивает большую достоверность в выводах, сделанных о физиологическом состоянии ткани. Во-вторых, определение относительного количества (или концентрации) двух основных сократительных белков позволяет экстраполировать трехмерные свойства ткани. Альтернативно заявлено, что если относительное содержание тяжелой цепи миозина и актина уменьшается с сопутствующим увеличением площади ткани или fCSA во время применения силовой тренировки , то это может отражать увеличение объема саркоплазмы . В-третьих,было показано, что этот метод обладает хорошей воспроизводимостью и чувствительностью (Roberts et al., 2018). Однако (!), основным ограничением метода является то, что данный анализ ткани проводят непосредственно после программы силовой тренировки (!), и поэтому возможное снижение относительного содержания тяжелой цепи миозина и актина по сути может говорить об локализованных отеках, а не о саркоплазматической гипертрофии.
Аналогичным образом, этот метод генерирует бинарную классификацию белков как миофибриллярных или саркоплазматических. Действительно, данные протеомики показали, что большинство миофибриллярных белков, полученных этим методом, являются сократительными или другими белками, связанными с миофибриллами, в то время как большинство саркоплазматических белков являются ферментами, которые находятся в саркоплазме (Vann et al., 2020b). Однако в каждой фракции содержатся ничтожные количества белков,которые можно считать загрязнителями. Как уже упоминалось выше, идентифицирован ядерный белок в миофибриллярной фракции, и белки сарколеммы были обнаружены в обеих фракциях. Было также обнаружено следовые количества изоформ ламинина и коллагена во фракции миофибрилл. Хотя это не является большим ограничением, исследователь должен также убедиться, что мышечная ткань явно свободна от крови, жира и соединительной ткани сразу после биопсии и до быстрого замораживания. В связи с этим ткани, замороженные с остаточной кровью, будут искусственно поднимать влажные мышечные массы, и эта техническая ошибка приведет к ложному заключению того, что относительное содержание сократительных белков ниже по сравнению с образцами тканей, которые свободны от крови. Наконец, суспензия миофибрилл в буфере 2 (описанная выше) представляет собой техническую проблему. В то время как имеется разработанная методика, которая солюбилизирует миофибриллы (Roberts et al., 2020) исследователи, заинтересованные в использовании этого метода, должны обеспечить надлежащую солюбилизацию, оптимизированную до масштабирования экспериментов.
4)Метод номер 4 - Оценка специфического напряжения в отдельных мышечных волокнах.
Этот метод выполняется с использованием специализированного датчика измерения напряжения, который может обеспечить косвенную оценку объема сократительного материала внутри волокна относительно его поперечному сечению (D'Antona et al., 2007; Degens et al., 2010; Meijer et al., 2015). Этот метод сначала включает в себя получение биопсии, криоконсервирование ткани и замораживание ее до дня эксперимента. В день эксперимента ткань размораживают и обрабатывают в растворе, содержащем “Тритон х-100”. Затем ткань помещают в расслабляющий раствор, отдельные волокна рассекают (или снимают кожу), а отдельное волокно подвешивают к противоположным штифтам нейлоновыми швами, которые взаимодействуют с датчиком напряжения. Напряжение волокна регулируется в расслабляющем растворе под микроскопом для достижения оптимальной длины саркомера (≈2,6 мкм). В этот момент диаметры волокон могут быть получены для экстраполяции значений площади поперечного сечения волокна (fCSA) . Затем волокна переносятся в физиологический раствор (рН 7,0), содержащий кальций, АТФ и другие компоненты. Данный раствор действует как активатор волокна .Изометрическая сила, развиваемая волокном, затем контролируется до тех пор, пока не будет достигнуто плато в развитии силы, и это значение может быть разделено на площадь поперечного сечения волокна для получения нормированного по площади волокна напряжения (т. е. удельного напряжения). В качестве дополнительного шага исследователи могут определить тип волокна, удалив волокно из устройства датчика силы, гомогенизируя его в определенных буферах, выполняя “SDS-PAGE с окрашиванием Кумассие” или серебром , наблюдая за рисунком изотипа тяжелой цепи миозина. Если выполнить этот метод с двумя волокнами, имеющими одинаковые диаметры, то можно справедливо заключить, что волокно, имеющее меньшую величину удельного натяжения, обладает меньшим количеством миофибрилл в поперечном относительно своего поперечного сечения (!). Сила данного метода заключается в функциональных данных, полученных на волокнах ex vivo.
В качестве контекстуального примера может быть сложно распутать роль нейронных адаптаций в адаптивном процессе, когда человек демонстрирует увеличение мышечной силы и гипертрофию в течение периода силовой тренировки. С этой целью анализ отдельных волокон может дать более четкие доказательства того, что гипертрофия мышечных волокон и, более конкретно, аккреция белков миофибрилл способствовали увеличению силы. Ограничением этого метода является то, что анализируются только сегменты отдельных волокон, а не волокна всей длины. Еще одним ограничением является то, что пермеабилизация изменяет внутренние характеристики волокон (!), такие как чувствительность к кальцию и осмотический баланс, и волокна искусственно расширяются из-за этих изменений (!), о чем свидетельствуют значительно большие значения fCSA по сравнению со значениями fCSA, полученными с помощью гистологии (!). Третье, и значительное, ограничение этой техники включает в себя деликатные и трудоемкие этапы, связанные с микродиссекциями, а также деликатную природу сопряжения волокон с преобразователем силы. Из-за работы, необходимой для выполнения этой методики, обычно анализируется менее 10 волокон на одного субъекта (Meijer et al., 2015). Наконец, как и в случае с ТЭМ, эта техника широко не применяется из-за необходимого специализированного оборудования и опыта.
Примечательно, что все методы, рассмотренные выше, требуют получения биопсий скелетных мышц от людей-участников, что само по себе может быть ограничением для многих лабораторий. Взятие биопсии Vastus lateralis также ограничено низкими выходами тканей (≈100 мг) относительно размера мышц. В связи с этим анализ трупов показывает, что масса всей латеральной мышцы vastus lateralis составляет в среднем ≈ 375 г у людей старше 80 лет (Ward et al., 2009), и эти массы, вероятно, в среднем превышают 500 г у более молодых, здоровых участников. Таким образом, биопсия могла бы взять только 1/500 часть одной четырехглавой мышцы. Эта цифра становится еще более ничтожной, если учесть, что ТЭМ анализирующий 1-2 мг ткани, исследования кожных волокон изучает ∼10 из сотен тысяч мышечных волокон vastus lateralis, гистологические исследования исследуют десятки волокон, а биохимические исследования изучают 20-30 мг ткани. Еще одним ограничением метода биопсии является то, что один и тот же пучок волокон предположительно не отбирается при выполнении метода в пред - и послеоперационные моменты времени.
Данные, которые поддерживают вызванную силовой тренировкой саркоплазматическую гипертрофию.
Макдугалл и соавторы. (1982) опубликовали первое исследование на людях, дающее доказательства того, что саркоплазматическая гипертрофия может значительно способствовать гипертрофии мышечных волокон. Примерно столетием ранее (Morpurgo, 1897) сообщил о результатах первого исследования гипертрофии, вызванной тренировкой, на животной модели, и автор полагал, что увеличение размера волокон в первую очередь связано с расширением не миофибриллярных компонентов . Более раннее сообщение о потенциальной саркоплазматической гипертрофии ,вызванной тренировкой было также отмечено у людей (Penman, 1969)., который использовал методы ТЭМ (Penman, 1969). Однако исследование Morpurgo проводилось на животных, которых тренировали на беговой дорожке, а исследование Пенмана было ограничено всего лишь 2-3 участниками, что исключало формальные статистические оценки. Макдугалл и др. (1982) обследовали пятерых участников, которые в течение 6 месяцев занимались силовой тренировкой разгибателей предплечий. Биопсия была получена из трехглавой мышцы до и после тренировочного вмешательства. Мышечные образцы анализировались на морфологию с использованием методов ТЭМ. Значения fCSA II типа увеличились в результате тренировки, и авторы дополнительно наблюдали уменьшение до 3% двумерного пространства, занятого миофибриллами (Р < 0,05), а также увеличение ∼15% пространства, занятого саркоплазмой (Р < 0,05). Более убедительными являются данные, приведенные в той же статье, от семи участников, которые имели многолетний стаж силовых тренировок. Эти люди обладали более крупными мышечными волокнами второго типа по сравнению с нетренированными участниками до 6-месячного тренировочного вмешательства, но также они показали на 30% более низкие значения объема, занятым миофибриллами, и в два раза большие значения занятые саркоплазмой. Toth et al. (2012) также использовали TEM, чтобы сообщить, что 18 недель тренировки уменьшили пространство, занимаемое миофибриллами ∼15% в мышечных волокнах vastus lateralis у здоровых людей. Мейер и др. (2015) впоследствии использовали методы изолированного волокна и сообщили, что бодибилдеры, которые обладали большими мышечными волокнами vastus lateralis, представили специфические значения напряжения, которые были на 40% ниже, чем у нетренированных участников. Наша группа использовала окрашивание SDS-PAGE и Coomassie, чтобы определить, что 6 недель очень высокообъемной тренировки уменьшили относительное содержание (на миллиграмм сухой ткани) тяжелой цепи миозина и актина на ∼30% в мышцах vastus lateralis у 15 хорошо тренированных участников (Haun et al., 2019b номер). Окрашивание фаллоидином также использовалось для того, чтобы показать снижение плотности белка актина у этих участников от пре - до после тренировочного периода.
Это исследование было ограничено, учитывая, что посттренировочная биопсия была собрана только через 24 часа после восстановления после последней тренировки. Несмотря на это, восемь из этих участников сделали третью биопсию через 8 дней после последнего тренировочного протокола, и признаки саркоплазматической гипертрофии все еще были заметны. В другом подобном исследовании на 15 ранее тренированных мужчинах студенческого возраста, биопсия vastus lateralis была взята до 10-недельного тренировочного вмешательства и через 72 ч после последнего тренировочного протокола(Vann et al., 2020а). По сравнению со всеми вышеупомянутыми исследованиями,эти участники выполняли тренировку с большим рабочим весом , где упражнения выполнялись с использованием 3-5 сетов по 3-8 повторений (с весом м∼80-90% от 1ПМ). В среднем значения значения fCSA II типа увеличивались на 19% (р < 0,05).В то время как окрашивание SDS-PAGE и Coomassie указывало на снижение относительного содержания тяжелых цепей миозина и белка актина на миллиграмм сухой ткани, всего на ∼3%, эти изменения до и после тренировки были статистически значимыми (Р < 0,05). Примечательно, что эти снижения сократительного белка на миллиграмм ткани были далеко не такими устойчивыми, как наблюдалось в предыдущем исследовании Haun et al. (2019b) (т. е. снижение на 30% ). Вместо гипертрофии волокон II типа, наблюдаемой во втором исследовании, эти данные предполагают, что заметная аккреция миофибриллярного белка, вероятно, произошла при краткосрочной силовой тренировке.
В совокупности семь исследований на людях, упомянутых выше, дают некоторые предпосылки того, что саркоплазматическая гипертрофия может возникать во время силовой тренировки . В то время как все исследования отличались в отношении продолжительности тренировки и схем нагрузки, только последнее исследование реализовало то, что обычно считается тренировкой с более высокой нагрузкой (т. е. выполнение менее пяти повторений за подход при >85% 1ПМ).
Доказательства опровергающие вызванную силовой тренировкой саркоплазматическую гипертрофию.
Прежде чем обсуждать данные на людях, которые опровергают саркоплазматическую гипертрофию, связанную с ростом волокон, важно обсудить ключевое исследование на грызунах, которое предшествовало исследованиям на человеке. Голдспинк в 1964 году опубликовал отчет, в котором участвовали четыре группы самок мышей. Две группы были обучены получать пищу, используя нагруженный сопротивлением шкивный аппарат, нагружающий двуглавую мышцу плеча. Две другие группы не были снабжены шкивными устройствами и служили контрольными группами. Через 25 дней эксперимента все мыши были умерщвлены, а бицепсы были гистологически исследованы с помощью световой микроскопии при увеличении в 1000 раз. Независимо от количества вводимой пищи, мыши, помещенные в систему шкивов, обладали волокнами, которые были на 30% больше по сравнению с волокнами контрольных животных,а также была отмечена сильная линейная зависимость между средней площадью поперечного сечения волокон (fCSA) и числом миофибрилл у мышей, размещенных в шкивных системах. Большие и предположительно гипертрофированные волокна у тренированных мышей также обладали меньшим пространством, занимаемым саркоплазмой по сравнению с более мелкими волокнами. Объясняя свои выводы, автор заявил (стр. 215):
"Показано, что количество миофибрилл на одно волокно линейно увеличивается с увеличением диаметра волокна. Это означает, что увеличение числа миофибрилл не происходит с той же скоростью, что и увеличение площади поперечного сечения волокна. Точно измерить диаметр миофибрилл с помощью световой микроскопии не удалось, но представляется наиболее вероятным, что они увеличиваются в объеме с увеличением размера волокна."
Хотя эти данные ограничиваются грызунами, они предполагают значительное количество миофибриллярного белка, которое сопровождается гипертрофией мышечных волокон, вызванной сопротивлением. Более того, данные Goldspink предполагают, что миофибриллы увеличиваются в объеме, а не в количестве, и также наблюдается небольшая степень упаковки миофибрилл – или непропорциональное увеличение аккреции белка миофибрилл относительно роста волокон ,которая может быть связана с вызванной нагрузкой гипертрофией волокон.
В то время как работа Голдспинка добавила огромные знания о поведении миофибрилл, ни одно человеческое исследование до сих пор не обеспечило такого уровня детализации. Однако в согласии с выводами Голдспинка существуют скудные человеческие данные, которые опровергают представление о том, что вызванная силовой тренировкой саркоплазматическая гипертрофия предшествует или способствует росту волокон или тканей. Люти и соавторы. (1986) использовали анализ TEM для изучения морфологических адаптаций в мышечных волокнах vastus lateralis у восьми нетренированных мужчин студенческого возраста, которые тренировались в течение 6 недель. В то время как площадь vastus lateralis увеличилась на 8,4%, значения fCSA и плотность миофибрилл остались неизменными. Эти данные свидетельствуют о том, что пропорциональное увеличение белка миофибрилл сопровождалось ростом мышц во время тренировок. Данные TEM также были объединены с данными компьютерной томографии середины бедра, чтобы оценить, что абсолютный объем, занимаемый миофибриллами,который увеличился на 10% по всей длине мышц-разгибателей ног. Критически важно то, что не было определено, произошло ли это увеличение содержания миофибрилл за счет создания новых миофибрилл, увеличения ранее существовавших миофибрилл и/или расщепления миофибрилл. Более того, предположение, что морфология, приобретенная с помощью TEM , применима ко всем мышцам разгибателей ног, имеет ограничения, учитывая, что TEM исследует чрезвычайно малое количество ткани, как обсуждалось выше. Наконец, отсутствие увеличения fCSA при тренировке вместо увеличения мышечной площади трудно согласовать и говорит о более широком вопросе отсутствия согласия между методами, используемыми для оценки гипертрофии. В последующем исследовании этой же исследовательской группы были изучены изображения TEM с высоким разрешением от тех же участников, чтобы определить изменения расстояния между толстыми и тонкими филаментами при тренировках (Claassen et al., 1989). Расстояние между миозиновыми нитями было одинаковым в моменты до и после тренировочного вмешательства. Эти результаты дают дополнительные доказательства того, что рост миофибриллярного белка был пропорционален росту тканей. Полученные ранее данные компьютерной томографии также были использованы в последнем отчете, чтобы показать, что рентгенологическая плотность мышц середины бедра увеличивается при тренировках. Интересно, что исследователи связали это последнее открытие с потенциальным увеличением упаковки миофибриллы. Хотя это вполне обоснованное заключение, оно также может быть связано со снижением внутримышечного содержания липидов или увеличением содержания не сократительных белков (например, белков внеклеточного матрикса). Кроме того, предположение авторов о том, что упаковка миофибрилл произошла, противоречит их предыдущим данным ТЭМ, предполагающим, что, хотя содержание миофибриллы увеличилось в абсолютном выражении, плотность миофибриллы и расстояние между ними были сохранены.
Исследование Trappe et al. (2000 год) исследовали специфические изменения напряжения в изолированных мышечных волокнах vastus lateralis у семи ранее нетренированных пожилых мужчин после 12 недель силовых тренировок. Хотя волокна типа I и IIa демонстрировали увеличение значений fCSA на 20% и 13%, удельные значения натяжения в обоих типах волокон были одинаковыми. Аналогично докладу Luthi et al. (1986), эти результаты предполагают пропорциональную аккрецию белка миофибрилл, сопровождаемую ростом волокон. Группа Trappe опубликовала последующее исследование на семи пожилых, ранее нетренированных женщин, которые участвовали в 12-недельных силовых тренировках (Trappe et al., 2001). В отличие от мужчин в их предыдущем отчете, волокна типа IIa не гипертрофировались. Кроме того, статистические сравнения показали, что конкретные значения напряжения в обоих типах волокон не были затронуты силовой тренировкой. Эти данные свидетельствуют о том, что тренировка не влияла на морфологию волокон типа IIa, а радиальный рост волокон типа I сопровождался пропорциональной аккрецией белка миофибрилл.
Аналогичные результаты были получены другими исследователями на шести ранее нетренированных мужчинах, прошедших 12-недельную силовую тренировку (Widrick et al., 2002); в частности, гипертрофия мышечных волокон vastus lateralis происходила без изменений в изолированных значениях специфического напряжения волокон.
Совсем группа ученых исследовала тканевые адаптации vastus lateralis, которые имели место у ранее нетренированных мужчин студенческого возраста, прошедших 12-недельную силовую тренировку (Roberts et al., 2018). Участники были разделены на группы с высоким и низким уровнем ответа на основе сводного балла изменений среднего fCSA до и после тренировочного вмешательства,двухэнергетической рентгеновской абсорбциометрии-определяемой общей массы мышечной ткани тела и толщины латеральной мышцы бедра. 13 респондентов с высоким уровнем ответа испытали увеличение средней fCSA на 34% и увеличение толщины вастус-латэралис 24%. Однако эти люди не испытывали никаких изменений в содержании тяжелых цепей миозина и белка актина на миллиграмм ткани, и то же самое было обнаружено для 12 других испытуемых, которые практически не проявляли изменений в среднем fCSA. Подобно многим исследованиям выше, наши результаты показывают, что рост белка миофибрилл был пропорционален росту волокон . В совокупности эти исследования в значительной степени предполагают, что рост мышечных волокон или тканей в ответ на 6-12 недель силовой тренировки происходит через обычную (миофибриллярную), а не саркоплазматическую гипертрофию. В связи с гипотезами, изложенными далее в обзоре, важно отметить, что во всех этих исследованиях рассматривались ранее неподготовленные участники.
Различные исследования также ставят под сомнение достоверность саркоплазматической гипертрофии. Например, Moore et al. (2009) сообщили, что одиа сессия силовых занятий вызывает значительное увеличение скорости синтеза миофибриллярного белка натощак и минимальные изменения скорости синтеза саркоплазматического белка в течение 5-часового периода после тренировки. Кроме того, эти авторы сообщили, что скорость синтеза саркоплазматического белка наиболее чувствительна к питательным веществам после физической нагрузки, а не к самой физической нагрузке. Гасье и др. (2012) опубликовали аналогичные результаты с использованием метода D2O tracer, чтобы показать, что одна сессия силовой работы для нижней части тела стимулировала увеличение скорости синтеза миофибриллярного белка,но влияла на общую скорость синтеза остальных белков в течение 16-часового периода после тренировки. Уилкинсон и др. (2014) также использовали метод трассировки D2O, чтобы показать, что 4 дня односторонней силовой тренировки для мышц ног в течение 8-дневного периода значительно увеличивали скорость синтеза миофибриллярного белка, не влияя на скорость синтеза саркоплазматического белка. Точно так же другие использовали метод трассировки D2O, чтобы показать, что скорость синтеза миофибриллярного белка повышается через несколько недель после начала силовой (Brook et al., 2015). Эти данные в совокупности предполагают, что миофибриллярный белковый пул, а не саркоплазматический белковый пул, может испытывать наиболее сильное расширение в начале тренировки.
Помимо этих краткосрочных исследований , существуют также долгосрочные исследования и перекрестные сравнения, которые законно ставят под сомнение, связана ли саркоплазматическая гипертрофия с долгосрочной мышечной гипертрофией. Alway et al. и соавт. (1988) также использовали анализ TEM, чтобы сообщить, что внутриклеточном пространстве, занимаемом миофибриллами в мышечных волокнах икроножной мышцы,которое было одинаковым между тренированными испытуемыми (силовая тренировка), тренированными выносливостью и сидячими субъектами, несмотря на то, что значения fCSA были самыми высокими в группе силовых тренировок субъектов. Shoepe и соавт. (2003) сравнили характеристики волокон vastus lateralis у шести нетренированных и шести хорошо тренированных мужчин студенческого возраста (опыт силовой работы 7 лет). В то время как значения fCSA I и II типов были значительно выше в хорошо тренированной группе, специфические значения напряжения были одинаковыми между группами, что наводило на мысль о длительной гипертрофии волокон с пропорциональной аккрецией белка миофибрилл. Два вышеупомянутых исследования показывают, что долгосрочный рост волокон при силовой тренировке происходит через обычную,а не саркоплазматическую гипертрофию. Однако эти данные противоречат другим данным. Пансараса и др. (2009) исследовали характеристики волокон vastus lateralis у пяти женщин студенческого возраста, которые тренировались в течение 1 года,где удельное натяжение значительно возросло в волокнах I и II типа, в то время как средние значения fCSA не демонстрировали никаких изменений от пре - до после тренировочного периода. Хотя эти результаты трудно согласовать, они подтверждают модель долгосрочной гипертрофии, когда упаковка миофибрилл происходила без увеличения гипертрофии мышечных волокон. Группа исследователей недавно исследовала толщину и характеристики тканей vastus lateralis у шести хорошо подготовленных мужчин студенческого возраста (средний возраст тренировки 10 лет) по сравнению с шестью нетренированными мужчинами (Vann et al., 2020b). В то время как толщина мышц была на 22% больше (Р < 0,05) в тренированной когорте, содержание тяжелых цепей миозина и актина на миллиграмм влажной мышечной ткани также было на 9% больше (Р < 0,05). Значения fCSA I и II типов были численно выше, но статистически схожи между когортами. D’Antona et al. (2006) также сообщили, что индивидуальные значения удельного натяжения волокон II типа были значительно выше у хорошо тренированных культуристов по сравнению с рекреационно тренированными контрольными группами. Опять же, результаты обоих исследований свидетельствуют о том, что длительная гипертрофия сопровождается большей упаковкой миофибрилл.
Выдвижение парадигмы с учетом коллективных доказательств
Учитывая вышеупомянутую литературу, трудно построить парадигму роста мышечных волокон, включающую в себя саркоплазматическую гипертрофию. Тем не менее, предполагается возможность трех сценариев, в которых саркоплазматическая гипертрофия может проявиться при силовой тренировке . В частности, предполагается, что саркоплазматическая гипертрофия является либо: (1) преходящим симптомом вызванного тренировкой отека, (2) преходящим механизмом роста мышечных волокон и/или (3) результатом достижения порога миофибриллярной аккреции белка у хорошо тренированных людей.
Отек тканей связан с локализованным повреждением и проявляется в виде задержки интерстициальной жидкости между клетками. Специфичный для вызванного физической нагрузкой повреждения мышц, отек может сопровождаться повреждением сарколеммы, когда инфильтрация крупных сывороточных белков (например, альбумина) и жидкости происходит внутри и вокруг мышечных волокон (Valle et al., 2013). В этих условиях представляется вероятным, что отек может искусственно повышать значения fCSA за счет задержки внутриклеточной жидкости, и это проявится как расширение саркоплазмы. В подтверждение этой гипотезы Yu et al. (2013) исследовали, насколько интенсивный бег вниз по лестнице влияет на среднюю подошвенной мышцы у мужчин студенческого возраста. Это исследование было ограничено тем, что повторные биопсии не были получены от участников; в частности, 16 мужчин выполнили тренировочный протокол, а биопсии были получены от участников через 1 час после тренировки, через 2-3 дня после тренировки и через 7-8 дней после тренировки. Седние значения fCSA были на 30% выше в пробах , полученных через 7-8 дней после физической нагрузки, по сравнению с пробами, полученными в более ранние моменты времени, и авторы предположили, что это было в первую очередь связано с локализованным отеком. Это исследование демонстрирует изменения fCSA через несколько дней после повреждения мышц из-за отека, и это, вероятно, проявляется через расширение саркоплазмы.
Неинвазивные оценки отека также могут быть выполнены путем захвата изображений мышц с помощью ультразвука или магнитно-резонансной томографии (МРТ) . Дамас и др. (2016c)отслеживали изменения vastus lateralis у нетренированных мужчин студенческого возраста до, через 3 недели и после 10-недельного тренировочного вмешательства. В то время как увеличение мышечной площади было зарегистрировано в 3-недельный и посттренировочный периоды времени, увеличение отека было обнаружено только в 3-недельный период времени. Авторы пришли к выводу, что гипертрофия мышц была смешана с отеком через 3 недели, но не через 10 недель после тренировки. Эта работа также привела к дискуссиям о том, можно ли точно оценить гипертрофию мышц без смешивающих эффектов отека в течение первого месяца тренировок у ранее нетренированных участников (Damas et al., 2016b; Defreitas et al., 2016). При соотнесении этих результатов с понятием саркоплазматической гипертрофии, вызванной тренировкой, вполне вероятно, что это явление может наблюдаться либо у нетренированных лиц в течение первого месяца тренировок, либо у тренированных лиц, подвергшихся воздействию непривычных тренировочных протоколов . Работа лаборатории Haun et al. (2019b), ,которая продемонстрировала облегченные тренировкой аспекты саркоплазматической гипертрофии, согласуется с последней гипотезой, поскольку участники, будучи хорошо тренированными, выполняли чрезвычайно высокие и непривычные объемы силовой работы . Еще одним подтверждением этого утверждения являются предварительные данные этих участников, показывающие, что увеличение содержания внеклеточной жидкости всего тела (оцениваемое с помощью биоэлектрической импедансной спектроскопии, или БИС) значительно способствовало увеличению изменений мышечной массы всего тела при тренировке (Haun et al., 2018). В то время как ранее предполагалось, что отек не смешивал переменные исхода в этом исследовании (Haun et al., 2019b), вышеупомянутые данные БИС действительно свидетельствуют об обратном.
Таким образом, продвигаясь вперед, исследователи должны учитывать смешивающие эффекты отека при измерении грубых суррогатов мышечной гипертрофии, а также морфологических или биохимических изменений в мышечной ткани. Кроме того, следует продолжать изучать влияние гидратационного статуса на макро - и микро - переменные, связанные с гипертрофией, учитывая, что во время хронических периодов силовой тренировки отмечались сдвиги жидкости (например, внутримышечная вода и объем плазмы) (Reidy et al., 2017).
Помимо того, что это потенциальный симптом отека, предполагается, что саркоплазматическая гипертрофия может быть механизмом, способствующим ранним фазам роста. Обычно средняя fCSA увеличивается на ∼15-30% в ответ на месяцы силовых тренировок (обзор в Grgic и Schoenfeld, 2018).Предполагается, что мышечные волокна должны в начале увеличить количество внутриклеточного пространства, чтобы в будущем накапливать больше мио фибриллярного белка . Одним из потенциальных механизмов достижения такого роста на начальном этапе может быть саркоплазматическая гипертрофия с последующим "обратным заполнением" вновь образованного пространства саркоплазмы миофибриллярным белком. Этот механизм противоречит концепции, выдвинутой Филлипсом (Phillips, 2000), где упаковка миофибриллы предшествует гипертрофии, а недавно добавленный материал миофибриллы “выталкивает” сарколемму наружу. Таким образом, мышечные волокна могут "расти" через саркоплазматическую гипертрофию в самом начале тренировочного вмешательства . Эта концепция далее проиллюстрирована на Рис.4.
В связи с рисунком 4, предложенная нами гипотеза расширения саркоплазмы гипертрофии мышц во время силовых тренировок включает в себя следующее:Наконец, предполагается, что третий сценарий, связанный с саркоплазматической гипертрофией, может включать "порог" аккреции белка миофибрилл, достигаемый в больших мышечных волокнах с годами тренировок. Опять же, эта гипотеза (как и две другие) является спекулятивной, учитывая, что, как и в случае с двумя другими вышеупомянутыми гипотезами, практически нет тому подтверждающих данных.
Ps: кому интересно читайте оригинал исследования https://www.frontiersin.org/articles/10.3389/fphys.2020.00816/full#B42