Сборка сообществ объясняет экологическую структуру морских сообществ в геологическом разрезе. Области с похожей историей диверсификации ("Эволюционные фауны") демонстрируют схожие результаты.
Расселение фауны на суше и в воде

Крупномасштабные модели расселения: члены этих фаун собрались на суше в начале палеозоя, затем распространились в море к концу палеозоя.
Эти модели существуют, несмотря на тот факт, что эволюционные фауны состоят из разрозненных как в филогенетическом, так и в экологическом отношении групп. Ранние работы показали, что схема сухопутной сборки и распространения на шельфе сообществ могут быть объяснены градиентом вымирания с более высоким коэффициентом вымирания на берегу.
Области с высокой степенью вымирания были заменены областями с более низкой степенью вымирания, которые затем распространяются на шельф.
Важность дифференцированных ставок происхождения на местах и вымирание также подкрепляется недавним анализом консервативности ниш внутри родов в это время, но роль эволюции ниши через миграцию или экспансию остается неизвестной.
Постпалеозойские области демонстрируют склонность к появлению самых ранних видов на суше, которые затем распространилась на море.
Это говорит о том, что эволюционные новинки в одних условиях встречаются чаще, чем в других, и эту гипотезу можно проверить на протяжении длительного времени с использованием эволюционных деревьев для исправления возможных проблем с отбором проб.
Градиент широтного разнообразия
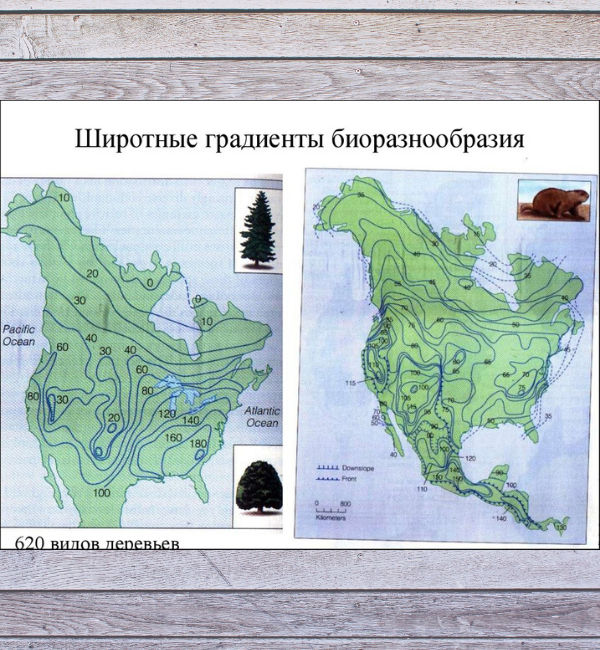
Филогенетическая палеоэкология также дает возможность проверить эту модель дифференциального поведения зарождения и вымирания. Градиент широтного разнообразия (LDG) - хорошо известная современная модель, имеющая глубокие эволюционные корни.
Исследования ископаемого запаса морских двустворчатых моллюсков показали, что современный ЛДГ обусловлен более высоким происхождением и более низким уровнем вымирания в тропиках по сравнению с внетропическими регионами, а также миграцией некоторых видов из тропиков.
Однако, этот современный ЛДГ возник всего 30 миллионов лет назад, и, возможно, не был постоянным с течением времени. Вместо этого, режимы ледников и тепличных хозяйств, как представляется, характеризуются различными моделями ЛДГ.
Решение вопроса о том, существуют ли эволюционные движущие силы для формирования ЦУР, и являются ли эти механизмы последовательными независимо от широты расположения для достижения пика разнообразия, требуется сочетание пространственно-временных моделей происхождения и исчезновение окаменелостей с филогенетическими связями вымерших таксонов.
Эволюция градиентов разнообразия во времени
Сохраняется много вопросов относительно эволюции градиентов разнообразия во времени, которые требуют знания эволюционных отношений.
Например, как скорость эволюции персонажей варьируются в зависимости от уклонов окружающей среды? Происходит ли расширение оффшорных зон главным образом за счет оккупации и диверсификации новых месторождений?
Учитывая, что эволюционных деревьев до сих пор не хватает для большинства морских обитателей с обширным послужным списком ископаемого, общинная палеоэкология может дать много полезной информации о происхождении и поддержании градиентов разнообразия по мере того, как эволюционные деревья становятся все более распространенными.
Избирательность при вымирании и облучении
Массовые вымирания видов и их восстановление могут оказывать несоразмерное воздействие на конкретные экологии, наблюдение, подкрепленное анализом как ископаемых, так и современных данных, включая филогенетические гипотезы, использованные в этих исследованиях.
Они позволяют различать, являются ли эти экологические сдвиги результатом случайных или селективных процессов.
Учитывая современную озабоченность по поводу современного биотического кризиса и массового вымирания, именно этот аспект филогенетической палеоэкологии имеет непосредственное отношение к биологии сохранения.