Если мы посмотрим на карту природных зон Земли (больше узнать о природных зонах Земли можно из тематических статей, ранее размещённых
на нашем канале:
то увидим, что они сменяют одна другую в определённой последовательности. На карте (на примере Северного полушария) это выглядит примерно следующим образом:
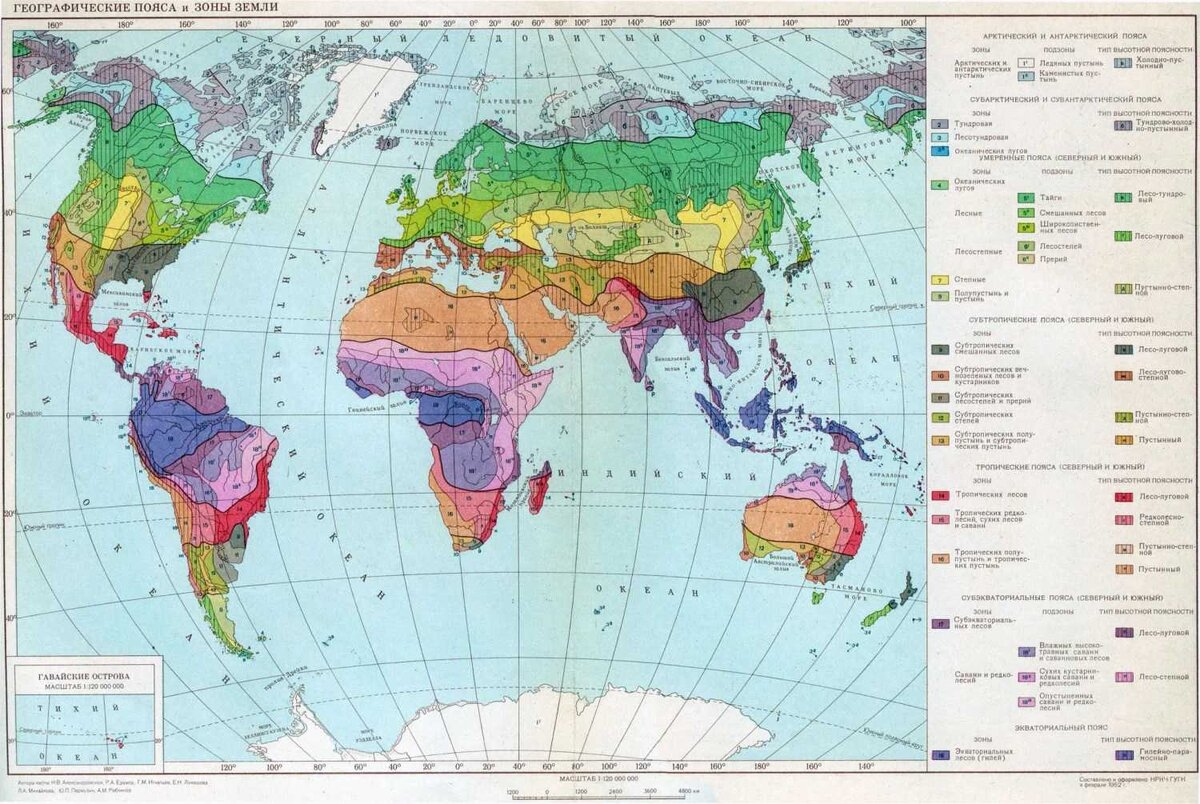
территории, располагающиеся в непосредственной близости от Северного полюса, занимает природная зона полярных пустынь, которая по мере удаления от полюса сменяется зоной тундр и лесотундр, на смену которой приходит зона лесов - сначала хвойных, затем смешанных, а потом и широколиственных, которые плавно переходят в лесостепи; лесостепи сменяются степями, степи - полупустынями, южнее которых располагаются уже настоящие пустыни. Дальше к югу пустыни сменяет зона саванн и редколесий - от примыкающих к пустыням и очень похожих на них опустыненных саванн до прилежащих с севера к зоне тропических лесов влажных лесосаванн, южнее которых начинается природная зона тропических лесов, которая простирается до самого экватора.
Что же служит причиной такой смены природных зон и соответствующих им типов растительности на нашей планете?
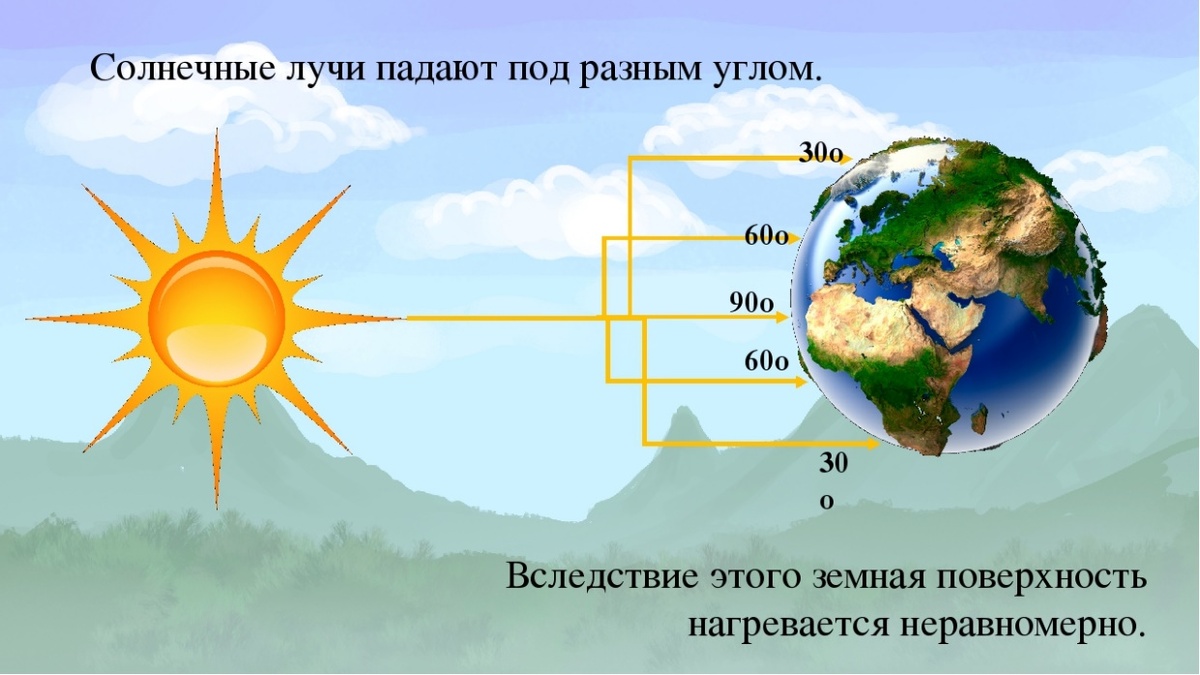
Таких причин несколько, но главных причин - всего две. Важнейшей из них является неравномерность распределения солнечной лучистой энергии на разных широтах, вызванная шарообразной формой нашей планеты и положением Земли относительно Солнца, вследствие чего солнечные лучи падают на земную поверхность под разными углами - тем меньшими, чем та или иная часть земной поверхности находится дальше от экватора - следовательно, чем дальше от экватора расположена определённая территория - тем меньше лучистой энергии Солнца она получает. Неравномерность распределения лучистой энергии Солнца по земной поверхности влияет на радиационный баланс каждой конкретной местности, следствием чего является неравномерное распределение тепла по поверхности Земли, что непосредственно влияет на циркуляцию атмосферы и влагооборот (больше узнать о циркуляции атмосферы можно из тематической статьи, ранее размещённой на нашем канале):
Второй по значимости причиной смены природных зон от полюсов к экватору является изменение соотношения тепла и влаги на той или иной широте, и именно этим обусловлено, например - наличие зоны лесов в субарктическом, умеренном, субтропическом, тропическом, субэкваториальном и экваториальном климатогеографических поясах, т.к. леса развиваются только на территориях, где осадков выпадает больше, чем может с этих территорий испариться влаги:
(больше узнать об осадках и о типах климата можно из тематических статей, ранее размещённых на нашем канале):
Каждая природная зона/экосистема подразделяется на несколько подзон/субэкосистем - биогеоценозов, при этом биогеоценозы могут сменять друг друга как в почти меридиональном направлении (пример: в природной зоне хвойных лесов в меридиональном направлении (с запада на восток) последовательно сменяются следующие биогеоценозы: ~ высокоствольные дождевые хвойные леса - состоят из хвойных деревьев многих видов - таких, как ель ситхинская, пихта Лоуа, туя гигантская, дугласия, кедр ладанный, кипарисовик Лавсона и др. (этот биогеоценоз представлен только на северо-западном побережье Северной Америки); ~ темнохвойные (елово-пихтовые) леса; ~ светлохвойные (сосновые) леса; ~ летнезелёные хвоепадные (лиственничные) леса), так и в широтном направлении (пример: в природной зоне степей и лесостепей в широтном направлении (с севера на юг) последовательно сменяются следующие биогеоценозы: ~ лесостепь; ~ кустарниковая степь; ~ высокотравно-разнотравная (луговая) степь; ~ дерновинно-злаковая (типичная) степь; ~ прутняково-эфемероидно-полынная (сухая) степь); смена биогеоценозов и в первом, и во втором случае обусловлена разницей в количестве выпадающих осадков, которое и определяет формирование каждого конкретного биогеоценоза на той или иной территории.
Для горных территорий широтная зональность, характерная для равнин - не действует, т.к. там имеет место т.н. высотная зональность, представляющая собой закономерную смену природных условий и, соответственно - природных зон/экосистем по мере возрастания высоты над уровнем моря. Это обусловлено тем, что с высотой возрастает разреженность воздуха, уменьшается атмосферное давление, происходит снижение температуры (подъём на 1 км даёт снижение температуры воздуха в среднем на 6°C), и т.д.. Высотная зональность лишь частично соответствует широтной зональности низменных территорий и включает значительно более узкий перечень природных зон (пример - высотная зональность южных склонов в Гималаях (от подножия к вершинам): ~ влажные тропические леса занимают высоты от 0 м над уровнем моря (далее - у.м.) до 1 600 м над у.м.; ~ вечнозелёные субтропические леса растут на высотах от 1 600 м до 2 500 м над у.м.; ~ широколиственные листопадные леса распространены на высотах от 2 500 м до 3 500 м над у.м.; ~ хвойные леса занимают высоты от 3 500 м до 4 000 м над у.м.; ~ субальпийское низкорослое криволесье располагается на высотах от 4 000 м до 4 300 м над у.м.; ~ альпийские горные луга покрывают горные склоны на высоте от 4 300 м до 4 800 м над у.м.; ~ высокогорная тундра (моховые болота и каменистые пустоши) располагается на высотах от 4 800 м до 5 300 м над у.м.; ~ нивально-гляциальная зона (зона вечных снегов и ледников, курумов и каменных осыпей) находится на высотах более 5 300 м над у.м. (больше узнать о ледниках и о криосфере Земли можно из тематической статьи, ранее размещённой на нашем канале):
Только в Гималаях из-за сочетания их месторасположения в тропическом поясе с их огромной высотой наблюдается такое количество высотных природных зон; во всех других горных системах, включая Анды в Южной Америке, количество зон высотной поясности меньше.
Естественная растительность, характерная для той или иной природной зоны, называется зональной растительностью. Она занимает наиболее типичные для каждой из природных зон местообитания, а именно - ровные
или всхолмлённые возвышенно-равнинные, хорошо дренированные водораздельные плато с условиями теплообеспечения, увлажнения и
почвами, наиболее типичными для той или иной природной зоны. Такие типичные местообитания зональной растительности называются плакорами. Зональность растительного покрова выражается в преобладании на плакорах тех или иных его жизненных форм - мхов и лишайников, трав, кустарничков и полукустарников, настоящих кустарников, деревьев, что при достаточном увлажнении лимитируется только средней температурой самого тёплого месяца в году (например, для произрастания мхов и лишайников лимитирующей температурой является 0°C, для произрастания трав - +5°С, для произрастания кустарников - +7°С, а для произрастания древесной растительности - +10°С). Именно поэтому в каждой природной зоне преобладает характерная именно для неё зональная растительность - тундровая (в зоне тундр), таёжная (в зоне хвойных лесов), хвойно-мелколиственная (в зоне смешанных лесов), листопадно-широколиственная (в зоне листопадных широколиственных лесов), и т.д..
Как мы помним из школьного курса географии, на нашей планете существует 13 (тринадцать) климатических поясов - по шесть одинаковых в Северном и Южном полушариях плюс экваториальный пояс по обе стороны от экватора примерно между 5° с.ш. и 5° ю.ш.. И в каждом из 10 (десяти) некрайних климатических поясов наблюдается чередование безлесных и лесных природных зон/биомов, а именно:
а) субарктический пояс - для него характерно наличие безлесного биогеоценоза лесотундры и лесного биома - хвойных лесов;
б) умеренный пояс - характеризуется наличием как лесных биомов (хвойных лесов; смешанных лесов; широколиственных лесов), так и безлесных биомов (лугов и пустошей; лесостепей и степей; полупустынь и пустынь);
в) субтропический пояс - для него также характерно наличие как безлесных биомов, таких как различные типы редколесий (например - маквис, фригана, гаррига, томиллар, шибляк, чаппараль, квонган, скрэб, мали, финбос, и др.), степи и лесостепи (прерии, пампа), пустыни и полупустыни, так и различных лесных биогеоценозов в составе биома субтропических лесов;
г) тропический пояс - в нём имеются и безлесные (пустыни и полупустыни; саванны и редколесья), и лесные (различные биогеоценозы тропических лесов) биомы;
д) субэкваториальный пояс - здесь также имеются биогеоценозы парковых саванн и влажных лесосаванн (условно безлесные), и биогеоценозы лесов с различными типами увлажнения (лесные);
Что же касается 3 (трёх) крайних климатогеографических поясов - двух полярных (северного и южного) и экваториального - то они характеризуются неполным развитием зональности, т.е. - отсутствием чередования лесных и безлесных природных зон ввиду того, что ~ в полярных поясах нет лесных биомов (это обусловлено прежде всего тем, что, как указывалось выше, в полярных климатических поясах отсутствует период со среднемесячными температурами выше +10°C, что исключает возможность произрастания там древесной растительности), а есть только безлесные - полярные пустыни и тундры; ~ в экваториальном поясе нет безлесных биомов, т.к. количество выпадающих осадков на этих территориях значительно превышает испаряемость, что при высоких среднемесячных температурах в течение
всего года полностью исключает наличие безлесных биомов естественного происхождения.
Как мы уже знаем, лесные биомы развиваются только при условии превышения количества выпадающих осадков над испаряемостью; это не относится к северному и южному полярным климатогеографическим поясам, в которых отсутствие лесных биогеоценозов обусловлено не превышением испаряемости над количеством выпадающих осадков (наоборот, в полярных поясах осадков выпадает намного больше, чем может испариться, хотя их количество в абсолютном выражении невелико - 100 - 400 мм в год), а отсутствием там календарных периодов со средней температурой выше +10°С, по изотерме которой и проходит граница распространения древесной растительности.
Для всех остальных безлесных биомов Земли характерны следующие соотношения количества выпадающих осадков и испаряемости: ~ зоны распространения лесостепей, саванн и редколесий характеризуются тем, что там количество выпадающих осадков примерно соответствует тому количеству влаги, которое может испариться, с незначительными вариациями по годам в ту или другую сторону; ~ зоны распространения степной и полупустынной растительности характеризуются тем, что количество выпадающих там осадков значительно меньше того количества влаги, которое может испариться с этих территорий; ~ зоны аридных пустынь формируются в условиях, когда испаряемость намного превышает количество выпадающих осадков.
Каждый тип зональной растительности характеризуется также такими показателями, как соотношение подземной и надземной растительной массы (измеряется в тоннах на 1 гектар площади) и водный режим почв каждого из зональных биомов или биогеоценозов в составе того или иного биома (больше узнать о типах почв Земли можно из тематической статьи, ранее размещённой на нашем канале):
Например, для ~ зон полярных пустынь и арктических и антарктических тундр соотношение подземной и надземной растительной массы (далее - СПНРМ) составляет 1:3, или 0,35 т к 1,05 т (для зоны полярных пустынь) и 5:1, или 10 т к 2 т (для зоны тундр), а водный режим (далее - ВР) полярнопустынных и тундровых почв характеризуется как мерзлотный; ~ для биогеоценоза лесотундры СПНРМ составляет 1:2, ВР - мерзлотный; ~ для биома хвойных лесов СПНРМ составляет 1:3,5, или 70 т к
245 т, ВР - промывной, кроме биогеоценоза летнезелёных хвоепадных лиственничных лесов, для которого ВР характеризуется как мерзлотный; ~ биомы смешанных лесов и лиственных лесов характеризуются так: СПНРМ составляет 1:3,2, или 95 т к 304 т, ВР - промывной; ~ для биогеоценоза лесостепи СПНРМ составляет 1:2,8, ВР - периодически-промывной; ~ для биогеоценозов степей и прерий СПНРМ составляет 4:1, или 20 т к 5 т, ВР - непромывной; ~ для биогеоценоза полупустынь СПНРМ составляет 4,5:1, ВР - непромывной; ~ для пустынь СПНРМ составляет 5:1, или 5 т к 1 т, ВР - выпотной; ~ для биома саванн и редколесий СПНРМ в среднем составляет примерно 1:3 - 1:5, а ВР для биогеоценоза опустыненных саванн - непромывной, для биогеоценоза типичных саванн - периодически-промывной, для биогеоценозов парковых саванн и влажных лесосаванн - промывной; ~ для биома субтропических лесов СПНРМ варьирует от 1:4 до 1:6 для разных биогеоценозов, а ВР определяется как промывной, кроме биогеоценоза летнесухих лесов средиземноморского типа, где ВР является периодически-промывным; ~ для биома тропических, субэкваториальных и экваториальных лесов СПНРМ варьирует от 1:9, или 10 т к 90 т - для биогеоценоза муссонных листопадных сухих лесов, до 1:4, или 100 т к 400 т - для биогеоценоза дождевых вечнозелёных экваториальных лесов, а ВР - от периодически-промывного до интенсивно-промывного в этих же биогеоценозах соответственно.
Как я уже писал выше, зональная растительность занимает наиболее типичные для каждой из природных зон местообитания - т.н. плакоры, и в каждой из природных зон преобладает характерная именно для той или иной зоны зональная растительность - тундровая - в зоне тундр, степная - в зоне степей, и т.д.. Однако в пределах каждой природной зоны кроме преобладающей там зональной растительности имеются также растительные сообщества, не входящие в её состав. Это т.н. азональная растительность.
Азональная растительность - это естественная растительность неплакорных местообитаний, встречающаяся в каждой из природных зон и не входящая в состав их зональной растительности. Азональная растительность включает в себя два типа растительных сообществ, а именно:
1. Экстразональная растительность - это растительность, являющаяся зональной для той или иной природной зоны, но произрастающая в пределах других природных зон, например: ~ участки степной растительности на южных склонах холмов в лесной зоне; ~ пойменные галерейные леса в безлесных зонах тундр, степей, саванн и пустынь; и т.д.;
2. Интразональная растительность - это такие биогеоценозы, которые ни в одной из природных зон/биомов не выходят на плакоры и нигде не образуют собственной отдельной природной зоны/биома; как правило, в своём распространении они связаны с такими местообитаниями, в которых условия водообеспеченности сильно отличаются от таковых на плакорах, например: ~ пойменные луга; болота; марши; солончаки; мангры; фитотельматы, и т.д.;
На фитотельматах хотелось бы остановиться особо.
Фитотельматы - это интразональные водно-болотные пресноводные
микроэкосистемы, распространённые во всех климатических поясах Земли, кроме полярных; представляют собой заполненные дождевой водой различные полости живых и/или мёртвых растений, в которых совместно обитают разнообразные живые организмы (бактерии, гетеротрофные протисты (больше узнать о типах питания живых организмов можно из тематической статьи, ранее размещённой на нашем канале):
микроскопические грибы, водоросли, черви, моллюски, ракообразные, насекомые, паукообразные, амфибии, некоторые виды млекопитающих).
Все фитотельматы на основании способа их формирования можно разделить на несколько типов:
А) Дендротельматы - это заполненные водой полости внутри дерева (дупла) или на его поверхности (углубления в развилках стволов и/или крупных ветвей); объём воды, находящейся в дендротельматах, может составлять от нескольких миллилитров до десятков литров;
Б) Стебли и междоузлия стеблей бамбука - образуют фитотельматы в результате механических повреждений или повреждений от деятельности насекомых-вредителей стеблей бамбука, в которых благодаря водонепроницаемости перегородок в междоузлиях скапливается дождевая вода, объём которой может достигать 1 л и более;
В) Листья, прицветники или лепестки растений - такие фитотельматы образуются в основном у однодольных растений из нескольких семейств (коммелиновые, лилейные, пальмовые, банановые, имбирные, геликониевые, пандановые, и др.), а также у двудольных из семейств зонтичные и жимолостные; могут содержать до 1 л воды;
Г) Ловчие кувшины хищных растений - это наполненные жидкостью (смесью дождевой воды и пищеварительных ферментов растения) ловчие органы насекомоядных растений из семейств Cephalotaceae (распространены в юго-западной части Австралии), Nepenthaceae (распространены от о. Мадагаскар на западе через Сейшельские о-ва, о. Шри-Ланка и тропическую часть Юго-Восточной Азии до северо-восточной части Австралии, о. Новая Гвинея и о. Новая Каледония), Sarraceniaceae (распространены в Северной и Южной Америке); ловчие кувшины некоторых видов непентесов могут содержать до 2 л жидкости;
Д) Резервуары для воды, расположенные в основании розеток листьев растений семейства Bromeliaceae/бромелиевых - могут располагаться как на земле, так и высоко над ней на стволах и в кронах деревьев (многие виды бромелиевых являются эпифитами и растут высоко над землёй); растения семейства бромелиевых распространены от юга США до центральной части Аргентины, а также на Малых и Больших Антильских о-вах и на небольшой территории в Западной Африке;
Фитотельматы представляют собой уникальные микроэкосистемы, предоставляющие широкие возможности для жизни, питания, укрытия, размножения многим видам беспозвоночных и позвоночных животных. Основу пищевых цепочек, формирующихся в фитотельматах, составляет детрит (детрит - это мёртвое органическое вещество, состоящее из мелких неразложившихся частиц останков различных растительных и животных организмов), образующийся при разложении органических веществ, а в ловчих кувшинах хищных растений - тонущие в них насекомые, разлагаемые пищеварительными ферментами этих растений и обитающими там бактериями. Многие виды животных не встречаются нигде, кроме фитотельматов. Помимо множества видов бактерий, простейших, микроскопических водорослей, грибов и других представителей микромира, в фитотельматах обитает также и много видов более крупных беспозвоночных животных - червей (плоских, круглых, кольчатых червей, пиявок), ракообразных (мелких веслоногих рачков, ракушковых рачков, ветвистоусых рачков, а также и крупных десятиногих ракообразных - например, таких, как сухопутный бромелиевый краб Metopaulias depressus, использующий бромелиевые фитотельматы для размножения, и т.д.), паукообразных (например, паук-птицеед Pachistopelma rufonigrum и паук-бокоход Thomisus nepenthiphilus живут только в фитотельматах, образуемых растениями семейства бромелиевых и семейства непентовых соответственно; при опасности эти пауки ныряют в воду, заполняющую фитотельмат), насекомых (ручейников, стрекоз, двукрылых, жесткокрылых, перепончатокрылых и т.д., которые используют фитотельматы в основном при размножении как места развития личинок, однако множество видов насекомых постоянно живут в фитотельматах и добывают там пищу, например, такие насекомые, как клоп-гребляк, стрекозы-стрелки, комары-звонцы, несколько видов жуков-плавунцов, трясинников и водолюбов, муравьи вида Colobopsis schmitzi, которые живут в ловчих кувшинах одного из видов непентеса и питаются насекомыми, попавшими туда, а также нектаром, выделяющимся по краям этих ловчих кувшинов для привлечения насекомых - потенциальных жертв; эти муравьи свободно плавают в толще и на поверхности жидкости, заполняющей ловчие кувшины, и не встречаются нигде, кроме этих фитотельматов). Кроме беспозвоночных, существует также достаточно много видов позвоночных животных, чья жизнь тесно связана с фитотельматами: это прежде всего более 100 видов лягушек 9 семейств, которые используют фитотельматы в качестве водоёмов для откладывания икры и последующего роста и развития вылупившихся из неё головастиков вплоть до превращения их во взрослых лягушек; обитающая в Юго-Восточной
Азии летучая мышь - гладконос Хардвика/Kerivoula hardwickii - также связана с одним из видов насекомоядных растений - Nepenthes hemsleyana - взаимовыгодными отношениями, а именно: летучие мыши используют ловчие кувшины непентеса в качестве укрытия и места для сна, а растение получает от летучих мышей их помёт, богатый азотом.
Образование, развитие и существование как зональных, так и азональных растительных сообществ всегда является результатом адаптации растений к условиям окружающей среды и отражает особенности таких компонентов ландшафта, как температурные условия, состав почв, особенности влагообеспечения, которые, в свою очередь, в той или иной степени зависят от рельефа местности.
Если вам понравилась статья - ставьте лайк, делитесь своими мыслями в комментариях и подписывайтесь на канал - будет интересно!
Если вы хотите поддержать автора, вы можете это сделать, нажав на ладонь с сердечком внизу справа. Спасибо!