Для любителей физиологии и биоэнергетики сегодня снова интересный материал. Речь пойдет о метаболической реакции на повторный спринт. Ученые смотрят изменения в гормонах и метаболитах крови и мышц.
Недавно у нас был похожий материал, где изучали механизмы энергообеспечения при работе руками:
Но, сегодня уже другое упражнение, продолжительность нагрузки и отдыха короче, плюс выполняется биопсия - есть возможность параллельно понаблюдать за изменениями в мышцах и в крови.
Идея исследования
Описать метаболические изменения при выполнении повторяющихся упражнения максимальной интенсивности.
Испытуемые и дизайн
8 здоровых физкультурников мужского пола (26,7±8,4 лет, 175,6±11,4 см, 71,8±11,4 кг) приняли участие в этом эксперименте.
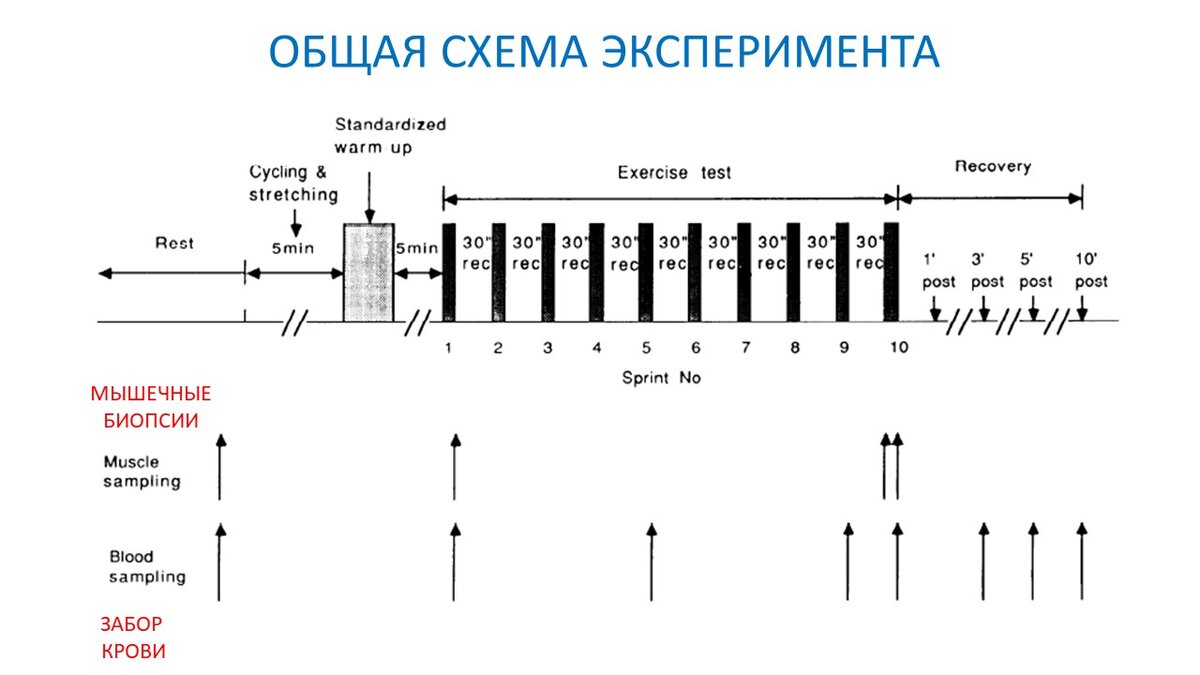
- Выполнялось десять 6-секундных максимальных спринтов на велоэргометре;
- Отдых между спринтами составлял 30 секунд;
- После последнего спринта испытуемые лежа восстанавливались в течение 10 минут (рис. 1);
Что измеряли
- Образцы крови брали в покое, сразу после первого, пятого, девятого и десятого спринта. А также на 3, 5 и 10 минуте восстановления (рис. 1);
- Биоптаты мышц брали в состоянии покоя, сразу после первого спринта из одной ноги и за 10 сек до и сразу после последнего спринта из другой ноги (рис. 1);
Результаты
Мощность
- Значения максимальной и средней мощности показаны на рис. 2;
- Максимальная мощность первого спринта (1253,3±334,8 Вт) была примерно в пять раз больше, чем мощность МПК при предварительном тестировании (253,6±57,8 Вт);
- В пятом и последнем спринте средняя мощность снизилась на 12,6% и 26,6% от максимального значения соответственно;
- Снижение пиковой мощности после 5 и 10 спринтов составило в среднем 15,9% и 33,4% соответственно;
Лактат
- После первого спринта наблюдалось небольшое повышение лактата в крови, однако после 5-го лактат увеличился в 15 раз с 0,6±0,1 до 9,2±1,5 ммоль/л (рис. 3);
- После завершения девятого спринта уровень лактата в крови увеличился до 12,6±1,5 ммоль/л и не изменился в течение 10 мин восстановления (рис. 3);
- Сильная корреляция (r = 0,88) обнаружена между увеличением лактата в крови в конце 5-го спринта и общим объемом работы, выполненной за первые 5 спринтов;
- Связь для вторых пяти спринтов, в ходе которых было выполнено 45,5% всей проделанной работы, не была такой сильной;
- Однако те люди, у которых наблюдалось наибольшее снижение средней мощности после 10 спринтов, как правило, имели самые большие изменения как в лактате крови через 5 минут после тренировки, так и в показателе pH крови сразу после 10 спринта;
- К концу первого спринта произошло лишь небольшое изменение показателя pH крови. Однако к концу упражнения показатель снизился на 0,27 единицы (с 7,37±0,03 до 7,10±0,03) и оставался низким в течение первых 3 мин восстановления (рис. 4);
- В течение последующих 7 мин восстановления показатель pH крови увеличился до 7,16±0,07, что указывает на то, что временной ход восстановления в этот период отличался от такового для лактата крови;
- Снижение концентрации ионов водорода в крови происходило быстрее, чем снижение лактата после тренировки;
Катехоламины плазмы (адреналин и норадреналин)
- Первый спринт привел к пятикратному увеличению концентрации адреналина в плазме с 0,4±0,2 до 1,9±1,0 нмоль/л и двукратному увеличению концентрации норадреналина с 1,7±0,4 до 3,3±1,1 нмоль/л (рис. 4);
- Концентрации адреналина и норадреналина в плазме увеличились после 5-го спринта до 4,2±1,4 и 15,7±2,7 нмоль/л соответственно;
- Пиковые значения наблюдались после девятого спринта, что представляет собой 13-кратное увеличение уровня адреналина до 5,1±1,5 нмоль/л и 13-кратное увеличение уровня норадреналина до 22,3±6,5 нмоль/л (рис. 4);
Глюкоза крови
Уровень глюкозы в крови увеличился с 3,9±0,2 до 4,6±0,5 ммоль/л после девяти спринтов и продолжал расти до 5 минут периода восстановления (5,6±0,6 ммоль/л);
Изменение объема плазмы
- Упражнение привело к среднему уменьшению объема плазмы на 12,1±4,5%;
- Изменения концентрации лактата, глюкозы и катехоламинов плазмы крови в ответ на нагрузку были в несколько раз выше, чем в объеме плазмы. Следовательно, изменения в метаболитах не могут быть объяснены изменениями объема плазмы;
Изменение мышечных метаболитов
- Концентрации мышечных метаболитов показаны на рисунке 5;
- После первого спринта произошло снижение концентрации креатинфосфата (КрФ) на 57%, что также сопровождалось снижением АТФ на 13%;
- Общая концентрация гликогена снизилась на 14% после 1-го спринта, тогда как наблюдалось резкое увеличение концентраций метаболитов гликолиза: глюкозо-6-фосфата, фруктозо-6-фосфата и концентрации лактата (рис. 5);
- Перед последним спринтом снижение концентрации АТФ на 32% сопровождалось снижением гликогена на 30% (рис. 5);
- Перед последним спринтом наблюдалось увеличение уровня глюкозы и лактата в мышцах по сравнению со значениями, наблюдавшимися после первого спринта;
- Хотя концентрация гликогена упала еще на 6% во время последнего спринта, не произошло почти никаких изменений в концентрации АТФ, промежуточных гликолитических метаболитов, глюкозы или лактата (рис. 5);
- Концентрация КрФ перед 10-м спринтом была на 51% ниже значения в состоянии покоя и упала до 16% во время последнего спринта;
Производство АТФ из анаэробных источников
- На долю гликолиза пришлось 44% АТФ, восстановленной во время первого спринта, а остальная часть была получена преимущественно в результате распада КрФ (рис. 6 и 8);
- Напротив, во время последнего спринта производство АТФ почти полностью обеспечивалось за счет КрФ (80%);
- Несмотря на вклад гликолиза в выработку АТФ в последнем спринте в среднем 20% (рис. 6), следует отметить, что у четырех испытуемых он был оценен как нулевой;
- В первом спринте анаэробный гликогенолиз превысил гликолиз на 91%, при этом скорость общего распада гликогена превысила скорость анаэробного гликогенолиза почти на 65% (рис. 7);
- После 10 спринтов произошло 11-кратное снижение гликогенолиза и 8-кратное снижение гликолиза, хотя общий распад гликогена снизился лишь на 37% (рис. 7);
- Люди, у которых наблюдалось наибольшее падение средней мощности в течение десяти спринтов, имели наибольшую скорость гликолиза во время первого спринта;
- Несмотря на высокую отрицательную корреляцию между скоростями гликогенолиза между первым и последним спринтом (r=-0,84), те, кто достиг наибольшего объема работы в первом спринте, также выполнили больше работы в 10-м спринте (r=0,851);
Выводы
- Результаты показывают, что мощность в первом спринте поддерживалась за счет энергии, которая была получена в основном за счет креатинфосфата и анаэробного гликогенолиза;
- Однако в последнем спринте накопления лактата не наблюдалось, но мощность все еще составляла 73% от мощности первого спринта;
- На основании косвенных данных, свидетельствующих об ингибировании анаэробного гликогенолиза в конце нагрузки, можно предположить, что ресинтез АТФ (в последнем спринте) в основном происходит за счет креатинфосфата и аэробного метаболизма.
PS
В общем, есть над чем подумать. Биоэнергетические вклады, как мы смотрели в работах ранее, при неполном восстановлении меняются.
Также хочется вспомнить в про расход гликогена в разных мышечных волокнах. Материал об этом у нас есть (ниже по ссылке). Но, к сожалению, в этой работе об этом не говорится.
Получить доступ к неопубликованным лекциям, статьям и другим материалам, а также поддержать нашу работу можно на Boosty
Сейчас там опубликованы следующие материалы:
1. Конкурентный тренинг (лекция).
2. Весогонка в единоборствах (лекция).
3. Плиометрика в фитнесе (лекция).
4. Метаболические эффекты упражнений (доклад).
5. Зона жиросжигания - что мы о ней знаем (лекция).
6. Физиология аэробного и анаэробного порогов, МПК (лекция).
7. Все про Дроп-Сет (лекция).
8. Локальное жиросжигание (лекция).
9. Ишемический тренинг. Часть 1 и 2 (научный доклад).
10. Физподготовка в волейболе (три лекции).
11. Биоэнергетика спорта (обзорная лекция).
12. Классификация аэробных и анаэробных упражнений (лекция).
13. Статодинамика: 30 лет обсуждений Часть 1 (лекция).
14. Физиология жиросжигания. Часть 1 и 2. (лекции по 3 часа).
15. NEW! Разное количество повторений. Обзор эффектов для массы, силы и выносливости (лекция).
16. NEW! Предпосылки для локального жиросжигания (доклад).
Дополнительные материалы по этой теме:
ТАБАТА: аэробная или анаэробная тренировка?
Анаэробной зоны в длительном беге не существует. Научный эксперимент
Экономят ли мышечный гликоген углеводные напитки? Научный эксперимент
Какая тренировка сильнее ускоряет метаболизм: силовая или интервальная?
Источник: Gaitanos GC, Williams C, Boobis LH, Brooks S. Human muscle metabolism during intermittent maximal exercise. J Appl Physiol (1985). 1993 Aug;75(2):712-9.