Это первая часть конспекта видеолекции Павла Пашковского "ВЛИЯНИЕ КАЧЕСТВА СВЕТА НА РАСТЕНИЯ РОЛЬ ОСНОВНЫХ ФОТОРЕЦЕПТОРОВ И КОМПОНЕНТОВ ИХ СИГНАЛИНГА. PROlvl". Полный конспект с таймингом и сырыми данными ведётся здесь. Слова и изображения взяты из лекции. Источники - в конце.
Я расскажу о рецепторах и пигментах. Фотосинтетические пигменты, как правило, локализованы в хлоропластах, участвуют в фотосинтезе, довольно стабильны. Рецепторы находятся в цитоплазме клетки, они отправляют сигнал в ядро, где активируется экспрессия соответствующих генов. Вот этот световой сигналинг идёт параллельно фотосинтезу, но очень сильно на него влияет. Сигналинг и фотосинтез взаимосвязаны.
Несколько слов о световом сигналинге
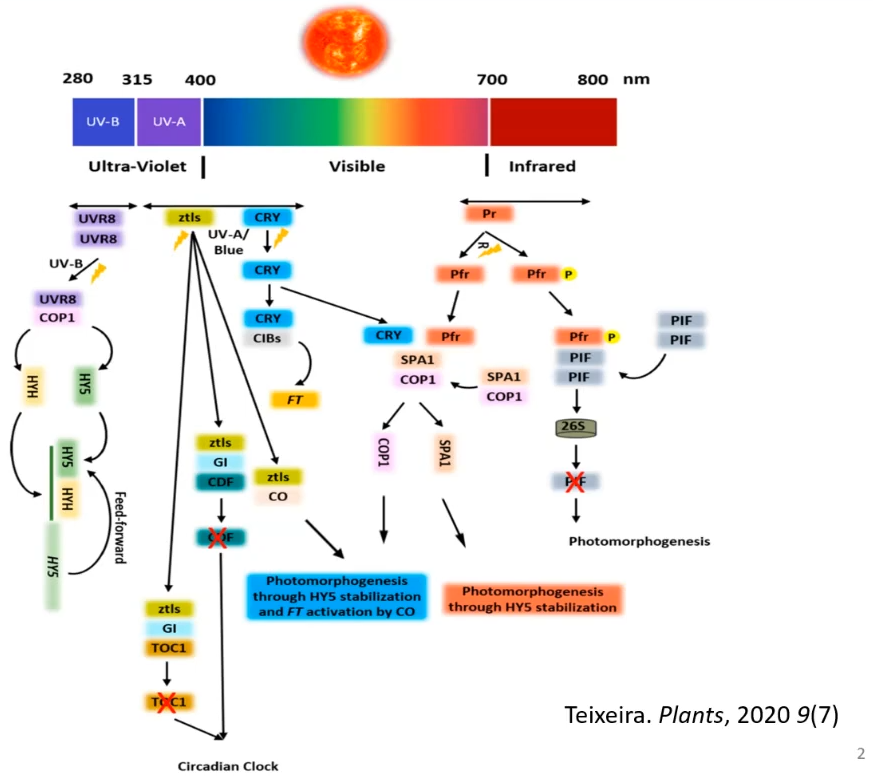
Качество света - очень важный параметр внешней среды, он оказывает решающее значение на большинство процессов, протекающих в клетках растений. Для определения качества света, его интенсивности, направленности в ходе эволюции у растений сформировалась сеть фоторецепторов, которые представляют собой специализированные белки, играющие решающую роль в большинстве процессов. Структура этих белков обычно состоит из хромофора (часть белковой молекулы, которая может возбуждаться тем или иным квантом света) и белковой части (которая и превращает сигнал в ответ растений на клеточном уровне).
На картинке выше представлены основные фоторецепторы:
- UV-B - рецепторы ультрафиолета B
- рецепторы, воспринимающие очень узкий световой диапазон
- криптохромы и зейтлуп (так называемые рецептороподобные, молекулы которых имеют в своей структуре компоненты, чувствительные к свету)
- фитохромы, отвечающие за рецепцию красной и дальней красной составляющей спектра
Тезисно о каждом фоторецепторе
Фитохромы
Фитохромы - очень важные в жизни растений фоторецепторы. Помимо непосредственно чувствительности к красной и дальней красной области спектра они выполняют функцию терморецепторов и также связаны с реакцией растений на стрессорные воздействия и способны к обратимости реакций. Фитохромы бывают двух типов:
- фитохромы типа A - считаются рецепторами I-го типа
- фитохром типа B - считаются рецепторами II-го типа
Фитохромы A
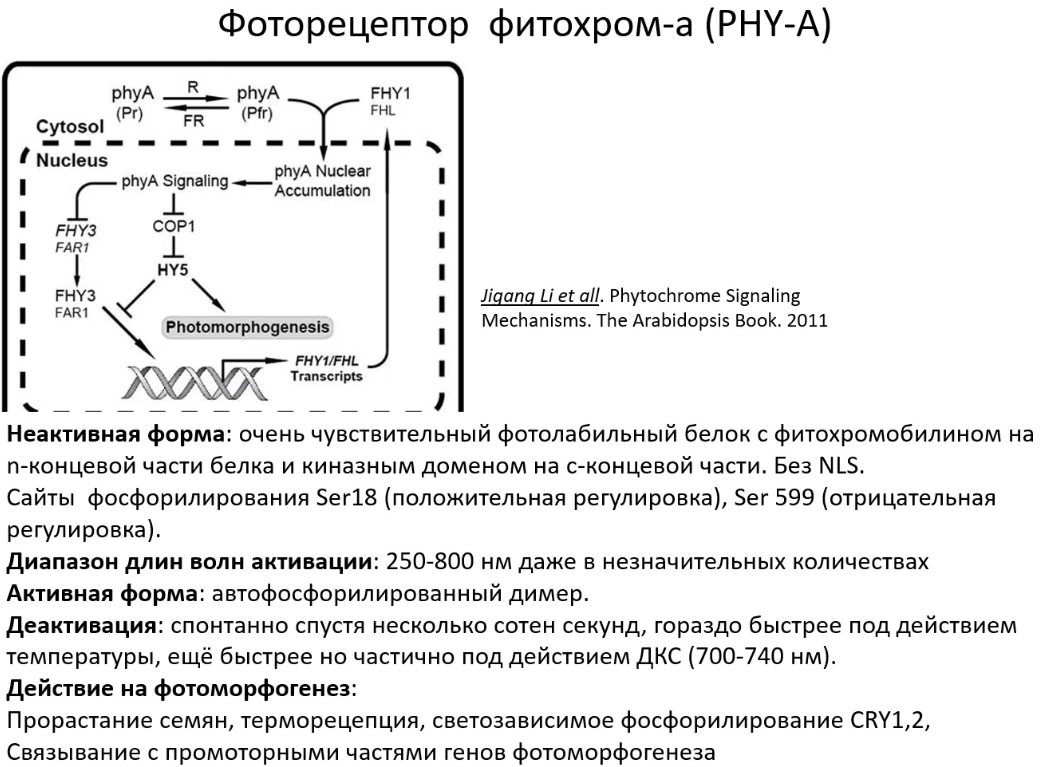
Их отличия в том, что фитохромы A, как правило, имеют необратимую реакцию и после рецепции света разрушаются. Но зато они выполняют очень важную роль - рецепцию как дальнего красного, так и инициацию светового сигналинга. Это происходит, например, в корнях прорастающих семян: семена ещё находятся в кожуре, а корень уже должен отправлять сигнал в семядоли о том, в какого качества света находится прорастающее растение. В этом как раз и выполняется функция фитохрома A.
Компонентами светового сигналинга являются несколько важных белков: белки транскрипционного фактора (HY5, COP1).
Транскрипционные факторы - это специализированные белки, которые способны связываться с промоутерными областями других генов и изменять их экспрессию. Промоутерная часть гена - это та часть гена, которая не участвует непосредственно в образовании белка, но с ней могут взаимодействовать компоненты, необходимые для синтеза белка, а также регуляторные компоненты. Так вот транскрипционные факторы - это и есть механизм регуляции этих генов. Можно сказать проще: гены-хозяева (транскрипционный фактор) управляют генами-рабами
Транскрипционные факторы типа HY5 - это "ключ ключей". У фоторецепторов много что различается (об этом позже), но есть что-то общее, и вот взаимодействие в HY5 является как раз одним из таких общих моментов, которые практически присутствуют у всех фоторецепторов. Также есть и другие транс. факторы.
Транскрипционные факторы типа HY5 являются позитивными факторами фотоморфогенеза, а убиквитин-лигазный комплекс, к которому относятся такие белки как COP1, являются негативными регуляторами. Световой сигналинг большинства рецепторов сводится к тому, что через комплекс посредников и вторичных мессенджеров (например, кальций) нужно передать сигнал с рецепторов на последующие низлежащие белки, чтобы ингибировать негативные факторы - убиквитин-лигазные комплексы, и активировать позитивные.
Фитохромы B
Есть ещё фитохромы B, C, D и т. д., но у большинства цветковых растений присутсвует 2 вида фитохромов, которые участвуют в сигналинге красного и дальнего красного света. Фитохром B также вовлечён в реакцию красного и дального красного света, но он способен переходить в 2 различные формы: неактивное и активное состояние. В активном состоянии он также отправляется из цитоплазмы в ядро, где непосредственно выполняет свою функцию: связывается с компонентами сигналинга вторичными месседжерами, и после этого идёт сигнал на транскрипционный фактор, который и выполняет активацию или деактивацию светового сигналинга.
Также есть HY5, от которого мы никуда не уходим. Но в фитохромном сигналинге фитохрома II типа есть ещё очень важные белки, другие транскриптиционные факторы, называемые PIF - факторы взаимодействия с фитохромом. Без них сигнал проходить будет очень плохо. Факторов фитохромного сигналинга очень много: у Arabidopsis их порядка 8 и не исключено, что будут открыты ещё. Среди них есть как HY5 (положительный фотоморфогенез), а PIFы сами по себе разделяются на группы, среди которых есть как положительные, так и негативные регуляции светового сигналинга.
Криптохромы (CRY)
У них реакция немного другая: происходит окислительно-восстановительная реакция, за счёт чего участвующий в этом фермент активирует рецептор, который также отправляется в ядро и там вызывает соответствующую световую реакцию. Также есть транс. факторы HY5. Появляются CO - это белки, которые вовлечены в цветение. Когда мы говорим про рецепцию, мы сразу же говорим не только про фотосинтез, не только про фотоморфогенез, а также про цветение, про переход к генеративной стадии или от цветения к плодоношению и реакцию на продолжительность дня и ночи.
Фототропин (PHOT)
Фототропины - это тоже рецепторы синего света. Как и у криптохромов, у них есть домены, которые могут окисляться и тем самым обеспечивать световой сигналинг. Но они выполняют немного разные функции. У фототропинов функция связана с выполнением настийных движений, положительного геотропизма и в дополнение к этому это одна из важных ролей в движении устьичных щелей: т. е. фототропины вовлечены в транспорт ионов, которые происходят при смыкании/размыкании устьичной щели, а также в сигналинг, который с этим связан.
Источники и персоналии
1. Пашковский Павел, лектор конспектируемого видео, кандидат биологических наук, ведущий научный сотрудник лаборатории физиологических и молекулярных механизмов адаптации Института физиологии растений им. К. А. Тимирязева РАН. Специалист по фотоморфогенезу.