Довольно популярная в палеокомьюнити тема - мегалодон (Otodus megalodon) против левиафана мелвилла (Livyatan melvillei), кто из них сильнее? Несмотря на всю бессмысленность подобного рода споров, это гипотетическое противостояние имеет гораздо больше смысла, чем аналогичные сравнения между мегалодоном и мозазавром (Mosasaurus hoffmanni), банально из-за реального факта сосуществования этих гигантских морских хищников (левиафан существовал около 9,9-8,9 миллионов лет назад; lambert et al., 2017, отодус мегалодон - около 16-3,5 миллионов лет назад; Boessenecker et al., 2019) в период миоцена. В прочем, если подобные вещи все ещё интересуют обитателей рунета, то следует раз и навсегда закрыть этот вопрос.
1. Размеры противников
Это, пожалуй, самый часто обсуждаемый аспект во всех сравнениях такого рода. Буду честен: глобально, на исход схватки он практически никак не повлияет, если вспомнить про все имеющиеся адаптации у сталкиваемых хищников для охоты на крупную добычу.
Начну с левиафана. Длина крупнейших особей может быть оценена в 17,5 метров, а масса может достигать 57 тонн (Villafana, J.A & Rivadeneira, M.M., 2014). Упоминать про его невероятно огромный череп - практически 3 метра в длину (270 сантиметров у голотипа; lambert et al., 2017) и 1,9 метра в ширину (lambert et al., 2010) особого смысла также не имеет.
Перейдем к мегалодону. Лишний раз говорить о его систематическом положении я сейчас не буду (уж слишком часто поднималась эта тема), просто скажу по факту: полностью взрослый мегалодон мог быть заметно больше взрослого левиафана. Предполагаемая максимальная длина гигантской акулы составляла 16-20 метров, с массой 16-и метровой особи в 61,56 тонн (Cooper et al., 2022). Нормальный измерений массы 20-и метровых особей, к сожалению, нет. Wroe et al., 2008, использования массу 20,3 метровой особи Carcharodon (!) megalodon в 103,2 тонны, что впечатляет даже на фоне крупнейших современных китообразных. Однако, само использование названия таксона C. megalodon может указывать на расчет массы на основе пропорций большой белой акулы (Carcharodon carcharias), увеличенной версией которой тогда считали мегалодона, поэтому такие оценки можно смело считать неактуальными. Более адекватными были бы расчеты на основе пропорций гигантской акулы (Cetorhinus maximus), более близкой к мегалодону как анатомически, так и локомоторно.
Сперва может показаться, что превосходство в размерах дает мегалодону определенное преимущество: он минимум на три тонны тяжелее противника и не уступает ему в длине. Но не будем торопиться. Дело в том, что современные зубатые киты - косатки (Orcinus orca) способны в одиночку убивать крупных китообразных, в том числе и кашалотов (Physeter macrocephalus; Jefferson et al., 1991) и взрослых гигантских (Cetorhinus maximus) и китовых (Rhincodon typus) акул (Fertl et al., 1996) больше себя. Если вспомнить, что вымерших макрохищных кашалотов, в том числе левиафана, сравнивают O. orca в плане экологии и относительных способностей (lambert et al., 2010), то от них тоже следовало ожидать чего-то подобного. Современные акулы также способны нападать на китообразных, и зубатые киты не становятся исключением (например, Heithaus, M.R., 2001), однако отодус мегалодон - это отдельный случай, ибо этот гигантский хищник имеет ряд анатомических отличий от современных (исключая представителей рода Somniosus) активных акул, о которых я рассмотрю ниже.


2. Вооружение
O. megalodon, судя по всему, обладал крайне сильными челюстями. Wroe et al., 2008 оценили силу укуса 103,2 тонной особи в 93127 Ньютонов для передний зубов и в 182201 Ньютонов для задних зубов (9,4-18,5 тонн). Аналогичные результаты для 47,69 тонной особи составили 55522 Ньютонов и 108514 Ньютонов соответственно (5,6-11,06 тонн, что не так уж и много, для животного таких размеров). Однако, Rice et al., 2016, на основе составленной ими регрессии, рассчитали силу укуса 17-и метровой особи O. megalodon в 123876 Ньютонов (12,6 тонн), а для 20,3 метровой особи была рассчитана сила укуса в 179219 Ньютонов (18,2 тонн), что не сильно меняет общую картину. Подобной силы мегалодону вполне хватало, чтобы прокусывать кости морских млекопитающих (достаточно неплохо они разобраны здесь), оставляя на них внушительные отметины от зубов.
Промеров силы укуса левиафана мелвилла нет (имеются лишь расчеты для его родственника - Zygophyseter varolai; Peri et al., 2021), однако предполагается, что сила сжатия его челюстей могла оставить позади любого крупного плиозавра (Paul., 2022). Его передние зубы, выступающие вперед под углом 45 градусов, короткий и широкий рострум, обеспечивающий более высокую силу укуса и бОльшую устойчивость к боковым нагрузкам, позволяли левиафану удерживать крупную добычу, оставлять глубокие раны и вырывать куски плоти из жертвы, подобно современной косатке (lambert et al., 2010).
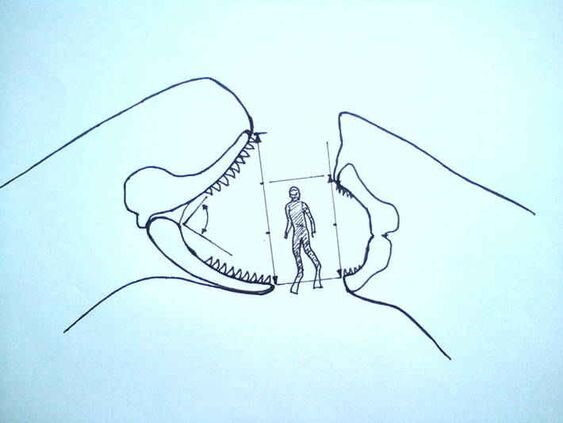
В целом, здесь можно признать примерное равенство: оба зверя обладают достаточно сильными челюстями, приспособленными к убийству крупной добычи. Кто-то может вспомнить про, якобы, "огромные трех и более метровые челюсти мегалодона". Здесь мне есть что ответить. На самом деле, зев пасти мегалодона был не таким большим, как многие себе представляют: он составлял всего лишь 1,2 метров при углу раскрытия пасти в 35 градусов и 1,8 метров при углу раскрытия пасти в 75 градусов соответственно, что может быть интерпретировано как адаптация к питанию относительно небольшими животными (Cooper et al., 2022). Что же касается L. melvillei, то расчетов максимального угла раскрытия его пасти я не нашел, однако, есть информация, на основе которой можно сделать какие либо выводы: современные P. macrocephalus способны разинуть пасть на впечатляющие 90 (!) градусов (Cantor et al., 2019). Если предположить, что левиафан мог открывать рот также широко, как современный кашалот, то он размер его раскрытых челюстей мог бы превысить аналогичные показатели у мегалодона, с учетом непропорционально огромного черепа вымершего китообразного. Также, некоторые могут вспомнить про невероятно большие зубы левиафана, однако, их длина в 36,2 сантиметров (самые большие зубы у голотипа) достигается за счет корня. Коронка зуба, выступающая из челюсти, была гораздо меньше (lambert et al., 2017).
3. Психология
Касательно мегалодона много сказать не получится: скорее всего, по всем своим поведенческим основам он был типичной акулой, просто очень больших размеров. Как нам известно, современные акулы не дерутся между собой. Да, могут проявлять "агрессию" по отношению друг к другу, однако. в большинстве случаев все ограничивается демонстрацией (по типу выгибания спины или мотания головой) или несильными укусами и ударами плавниками для изгнания чужака со своей территории или установления иерархии. Назвать "дракой" все эти проявления "агрессии" язык не поворачивается в любом случае, поскольку практически во всех случаях атакуемая сторона даже не пытается оказать сопротивление, за редкими исключениями (Klimley et al., 2023). Случаи хищничества по отношению к другим акулам я даже учитывать не буду, поскольку это типичное и ничем не примечательное пищевое поведение. Так что, как бы акулолюбы не кричали о "агрессивном и боевом отодусе мегалодоне", их фантазии не доказываются примерно... Ничем.
Левиафан мелвилла является членом надсемейства Physeteroidea. Его крупным современным родственникам - P. macrocephalus свойственны брачные сражения между самцами. В период размножения (когда животное достигает возраста в 20 лет) самцы кашалотов, возвращающиеся к местам размножения, как правило, никак не контактируют между собой, сводя контакт к нулю. Однако, если крупные самцы находятся в пределах одной группы между ними могут развязываться жестокие сражения, в ходе которых животные оставляют друг другу глубокие шрамы своими зубами (Whitehead., 2018). Судя по частоте наблюдения подобного рода повреждений у кашалотов разных возрастов, взрослые особи чаще проявляют агрессию по отношению друг к другу, чем молодь (Wursig, B & Orbach, D.N., 2023). Для современных O. orca, по всей видимости, норма наносить друг другу повреждения в брачный период или при коммуникации: многие самцы и самки несут на себе шрамы от зубов других косаток, а некоторые самцы получали травмы спинного плавника после таких схваток. Уровень внутривидовой агрессии O. orca может оказаться еще выше, если большинство сражений не оставляют на телах китообразных видимых следов (например, киты могли использовать свои рострумы и хвосты для нанесения ударов). Судя по всему, самки и подростки косаток реже калечат друг друга, чем взрослые самцы (Wursig, B & Orbach, D.N., 2023). Если экстраполировать поведение современных зубатых китов на L. melvillei, то можно предположить, что левиафаны также могли вступать в конфронтации с представителями своего вида, оставляя серьезные травмы на головах сородичей, подобно нынешним косаткам и кашалотам.
Отдельно хочется отметить один весьма интересный момент, связанный с взаимодействием современных нам акул и зубатых китов. Дело в том, что O. orca нередко включают самые разные виды акул в свой рацион. Сами же акулы опасаются крупных зубатых китообразных, причем настолько, что взрослые особи C. carcharias, после нескольких хищничеств косаток на них в водах Южной Африки, в страхе покинули свой регион обитания (Towner et al., 2022). Даже распиаренные акулофилами светящиеся акулы (Isistius), способные прогрызть обшивку подводной лодки (Фёдоров., 2017) и охотно поедающие китообразных, В том числе - зубатых, обходят взрослых косаток стороной (Feunteun et al., 2018). Интересно еще и то, что крупные акулы, несмотря на очевидную возможность убивать крупных дельфинов (как это было, например, с гриндами (Globicephala) и малыми косатками (Pseudorca crassidens)), не рискуют нападать на O. orca, в то время как большая белая акула для соразмерной ей самки косатки - это вполне неплохой источник пищи (Pyle et al., 1999). Вполне возможно, что при столкновении мегалодона и левиафана именно кит был бы нападающей стороной, ввиду неконфликтности гигантской рыбы и её специализации на питании относительно небольшими животными.
4. Другие факторы
Не вижу смысла что либо говорить об умственном превосходстве кита над акулой - слишком уж это очевидно. Преимущество левиафана также заключается в бОльшей скорости плавания, чем у мегалодона. Скорость плавания современных кашалотов, как показали измерения, может достигать 9,6 м/с, что эквивалентно приблизительно 34,56 км/ч при крейсерной скорости в 6,48 км/ч (Aoki et al., 2007) и, буду честен, говорить о меньшей скорости у L. melvillei, особенно, при меньших абсолютных размерах, у нас нет. Крейсерная скорость O. megalodon оценивается в 5,04 км/ч, с учетом возможности рыбы разгоняться до 37,15 км/ч при резких рывках, что даже с учетом его региональной эндотермии выглядит весьма скромно (Ferron., 2017). То есть, мы получаем достаточно быстрого и подвижного кита, способного длительное время поддерживать достаточно высокую скорость, и огромную медлительную акулу, способную лишь на непродолжительные скачки активности. Весьма неутешительный результат для фанатов мегалодона....
Возможно, что мегалодон имел бы неплохие шансы против левиафана в прямом столкновении, но никто не заставляет левиафана лезть на акулу-переростка в лобовую. Судя по повреждениям на позвонке мелкого миоценового усатого, атаковавший его хищник использовал таран для иммобилизации жертвы (такая тактика атаки не свойственна акулам, поэтому отодонтиды здесь отпадают автоматически). Современные косатки, как известно, часто таранят свою добычу: горбатых китов (Megaptera novaeangliae, что примечательно, одного прямого удара рострумом взрослой O. orca хватило, чтобы мгновенно убить огромного кита; Melnikov, V & Zagrebin, I.A., 2005) и морских слонов (Mirounga; Black et al., 2023), поэтому, я не вижу ничего, что мешало бы взрослому левиафану также протаранить мегалодона снизу, тем самым сломав ему позвоночник. Предел прочности акульих позвонков составляет приблизительно 25 МПа (Porter et al., 2006), а с учетом относительно небольшого размера позвонков O. megalodon - до 23 сантиметров в диаметре (Cooper et al., 2022), такой сценарий кажется вполне вероятным. Если же левиафан решит попробовать загрызть мегалодона, как косатка белую акулу, то уже тут он столкнется с проблемой в виде акульих плакоидных чешуек, о которые, со временем, стираются даже армированные зубы косаток (Ford et al., 2011). Однако, процесс это явно не быстрый, и стираться зубы будут постепенно, так что, вопреки фантазиям акулофанов, левиафан, убивая мегалодона своими челюстями, практически ничего не потеряет. Конечно, при полноценной специализации на питании мегалодонами эта проблема станет куда более значительной, но ни о какой специализации L. melvillei на питании акулами в научном сообществе и речи не идет (lambert et al., 2010)...
5. Результаты
Что-ж, пожалуй, все хоть немного важные аспекты были мной разобраны, так-что теперь можно сделать определенные выводы.
Так кто-же был королем морей миоцена - левиафана мелвилла или отодус мегалодон? Как мы видим, все рассмотренные мной детали указывают на то, что левиафан имел больше шансов победить в этом гипотетическом противостоянии за счет преимущества в скорости, интеллекте, общей прочности скелета и адаптаций к убийству и удержанию крупной добычи. Впрочем, неудивительно, что левиафану приписывают статус высшего хищника (lambert et al., 2010): для полностью взрослой особи L. melvillei, условный цетотерий, более мелкий зубатый кит и соразмерная ему (или более крупная) особь O. megalodon - это тоже самое, что для современной косатки усатый кит, дельфин и взрослая белая/гигантская акула соответственно =) Если вспомнить, как сильно C. carcharias боятся O. orca, то можно предположить, что левиафаны, чисто технически, также могли держать в страхе взрослых мегалодонов. И, к сожалению для акулофилов, всякие случаи хищничества мегалодона аж на мелких кашалотиков и кормление на трупе левиафана никак не меняет ситуацию...
Список литературы:
1) Lambert, O., Bianucci, G., De Muizon, C., Notes, A., 2017. Macroraptorial sperm whales (Cetacea, Odontoceti, Physeteroidea) from the Miocene of Peru. Zoological Journal of the Linnean Society 179.
2) Boessenecker, R.W., Ehret, D.J., Long, D.J., Churchill, M., Martin, E., Boessenecker, S.J., 2019. The Early Pliocene extinction of the mega-toothed shark Otodus megalodon: a view from the eastern North Pacific. PeerJ 7(2).
3) Villafana, J.A., Rivadeneira, M.M., 2014. Rise and fall in diversity of Neogene marine vertebrates on the temperate Pacific coast of South America Introduction. Paleobiology 40(4):659-674.
4) Cooper, J.A., Hutchinson, J.R., Bernvi, D.C., Cliff, G., Wilson, R.P., Dicken, M.L., Menzel, J., Wroe, S., Pirlo, J., Pimiento, C., 2022. The extinct shark Otodus megalodon was a transoceanic superpredator: Inferences from 3D modeling. Science Advances 8(33):eabm9424.
5) Wroe, S.W., Lowry, M., McHenry, C.R., Huber, D.R., Hutchinson, J.R., Cliff, G., Wilson, R.P., Dicken, M.L., Menzel, J., Cooper, J., Pirlo, J., Pimiento, C., 2008. Three-dimensional computer analysis of white shark jaw mechanics: How hard can a great white bite? Journal of Zoology 276(4):336-342.
6) Rice, K.W., Buchholz, R., Parsons, G.R., 2016. Correlates of bite force in the Atlantic sharpnose shark, Rhizoprionodon terraenovae. Marine Biology, 163(2).
7) Jefferson, T.A., Stacey, P.J., Baird, R.W., 1991. A review of Killer Whale interactions with other marine mammals: predation to co‐existence. Mammal Review 21(4):151 - 180.
8) Fertl, D., Acevedo-Gutierrez, A., Darby, F.L., 1996. A report of killer whales (Orcinus orca) feeding on a carcharhinid shark in Costa Rica. Marine Mammal Science 12(4).
9) Heithaus, M.R., 2001. Predator-prey and competitive interactions between sharks (Order Selachii) and dolphins (suborder Odontoceti): A review. Journal of Zoology 253(1).
10) Peri, F., Falkingham, P.L., Collareta, A., Bianucci, G., 2021. Biting in the Miocene seas: estimation of the bite force of the macroraptorial sperm whale Zygophyseter varolai using finite element analysis. Historical Biology 34(12).
11) Paul, G.S., 2022. The Princeton field guide to Mesozoic sea reptiles. Princeton University Press.
12) Cantor, M., Gero, S., Whitehead, H., Rendell, L., 2019. Sperm Whale: The Largest Toothed Creature on Earth. Ethology and Behavioral Ecology of Odontocetes (pp.261-280).
13) Klimley, A.P., Porcher, I.F., Clua, E., Pratt, H., 2023. A review of the behaviours of the Chondrichthyes: a multi-species ethogram for the chimaeras, sharks, and rays. Behaviour 160(11-14).
14) Whitehead, H., 2018. Sperm whale. Encyclopedia of Marine Mammals, 919–925.
15) Wursig, B., Orbach, D.N., 2023. Sex in Cetaceans Morphology, Behavior, and the Evolution of Sexual Strategies. Springer.
16) Towner, A., Watson, R., Kock, A.A., Papastamatiou, Y.P., Sturup, M., Gennari, E., Baker, K., Booth, T., Dicken, M., Chivell, W., Elwen, S., Kaschke, T., Ed Edwards, D., Smale, M.J., 2022. Fear at the top: killer whale predation drives white shark absence at South Africa’s largest aggregation site. African Journal of Marine Science 44(2):139-152.
17) Feunteun, A., De Schrevel, C., Verhaegen, M., Damien, C., Duchemin, M.B., Ziani, N., de Montgolfier, B., 2018. First evaluation of the Cookie-cutter sharks (Isistius sp.) predation pattern on different cetacean species in Martinique. Environmental Biology of Fishes 101(4):1-11.
18) Pyle, P., Schramm, M.J., Anderson, S.D., Keiper, C., 1999. Predation on a white shark (Carcharodon carcharias) by a killer whale (Orcinus orca) and a possible case of competitive displacement. Marine Mammal Science 15(2):563-568.
19) Фёдоров, К., 2017. Светящаяся акула. Тайны XX века.
20) Aoki, K., Amano, M., Sugiyama, N., Muramoto, H., 2007. Measurement of swimming speed in sperm whales. Underwater Technology and Workshop on Scientific Use of Submarine Cables and Related Technologies, 2007. Symposium on.
21) Ferron, H.G., 2017. Regional endothermy as a trigger for gigantism in some extinct macropredatory sharks. PLoS ONE 12(9):e0185185.
22) Melnikov, V., Zagrebin, I.A., 2005. Killer Whale predation in coastal waters of the Chukotka Peninsula. Marine Mammal Science 21(3):550-556.
23) Black, N.A., Talty, C.M., Schulman-Janiger, A., Srinivasan, M., 2023. Mammal Hunting Killer Whales off Monterey, California: A 30-Year Synthesis. Springer.
24) Porter, M.E., Beltran, J.L., Koob, T.J., Summers, A.P., 2006. Material properties and biochemical composition of mineralized vertebral cartilage in seven elasmobranch species (Chondrichthyes). Journal of Experimental Biology 209(Pt 15):2920-8.
25) Ford, J.K.B., Ellis, G., Matkin, C.O., Wetklo, M., 2011. Shark predation and tooth wear in a population of northeastern Pacific killer whales. Aquatic Biology 11(3):213-224.