Если вы следите за новостями, то наверно уже слышали об обнаружении и описании одной новой серобактерии. И это была бы обычная статейка, но бактерия эта длиной в два сантиметра. Да еще мало того, что она такая огромная, так она еще и полиплоидная, то есть содержит несколько кратных копий своего генотипа. Но мы сегодня не будем останавливаться на уже десятками людей обозначенной теме - оставим в покое эту бедную серобактерию на дне её мангровых зарослей.
Нет молодые люди, сегодня мы поговорим о такой страшной вещи как эукариотизация, то есть появление клеток с ядром и хромосомами, а также посмотрим, при чем здесь полиплоидность, археи и митоз. Будет страшно, обещаю! =)
NANOMACHINES SON!
Многие из нас, еще со школьной скамьи знают, что у бактерий есть только одна хромосома - та самая, что рисуют на схемах в виде колечка. Но на деле оказывается всё куда сложнее, и иногда таких колечек в клетке не одно, и не два, и не три. Сразу говорю - речь тут не о каких то дополнительных плазмидах, ни в коем случае. Я говорю действительно о том, что в одной клетке плавает сразу несколько полных копий генома.

Изначально полиплоидность для бактерий считалась чем-то редким и несерьезным - так, практически случайность. Но как оказалось, этим балуется большое количество прокариот - те же галобактерии или к примеру термоплазмы. Если сказать проще, то многие археи - группа прокариот близкая к эукариотам - имеют разные степени плоидности. Но зачем это им?
НАЧИНАЕМ УСЛОЖНЯТЬСЯ
На самом деле ответ довольно прост - такая система из нескольких копий генотипа нужна археям, чтобы выдерживать экстремальные условия, в которых они живут. К примеру та же термоплазма может спокойно сидеть в источнике воды с температурой в 55 градусов и рН равным 2. У этих бактерий сложная модифицированная мембрана, нет клеточной стенки, множество жгутиков. А главное - их ДНК образовывает комплексы с особыми белками, которые по строению гомологичны белкам-гистонам эукариот. Эти белки кстати помогают сохранять структуру ДНК и еще много для чего нужны и у обычных бактерий их нет, только у самых продвинутых архей. А еще такие белки есть у нас, у эукариот, хоть в гейзерах мы и не живем - ВНЕЗАПНО.
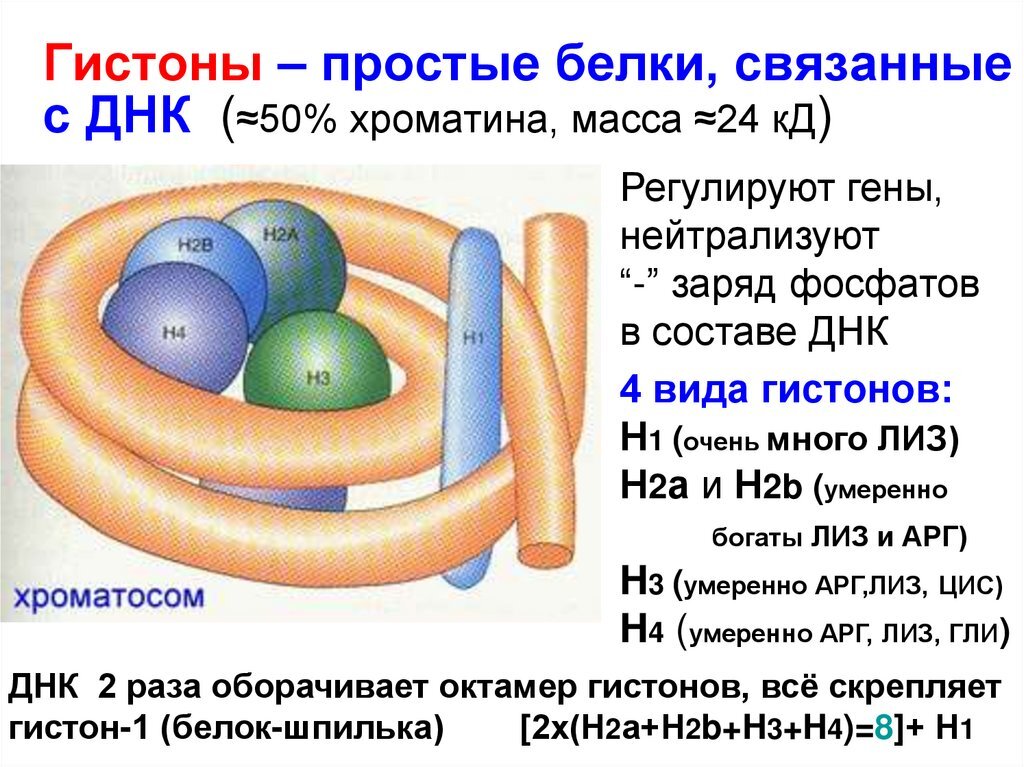
Так вот, несколько копий всего целиком генотипа позволяют по факту иметь несколько запасных копий любого гена, и в случае поломки этого гена в одной из хромосом всегда можно скопировать инфу с неповрежденного варианта. Не все конечно это умеют делать, но согласитесь, когда вы живете в очень плохих термически и химически условиях - лучше иметь запас генов для считывания, чтобы не загнуться. И конечно же научиться такой фишкой пользоваться.
Кроме того, стоит заметить, что при наличии нескольких одинаковых хромосом, такая вот полиплоидная архея очень хорошо защищена от множества рецессивных мутаций - ведь всегда есть в заначке рабочая доминантная версия гена. Удобно, ничего не скажешь. Кроме того, если у тебя настал внезапный дефицит фосфора, то ты всегда можешь уничтожить одну из хромосом. Вот вы думали что у ДНК есть только наследственная функция, а оказывается у некоторых архей она может использоваться также, как человек использует жир (почти). Чудеса в решете, да и только.
К тому же, оказывается, что при появлении полиплоидных клеток в бактериальной популяции, они оказываются куда успешнее, нежели клетки с одной хромосомой. Ну тут на самом деле срабатывают все описанные мною фишки разом - устойчивость к единичным генетическим мутациям здесь вообще является самой бомбической способностью полиплоидов. Правда это только первые несколько сотен поколений. Оказывается после своего эволюционного взлета полиплоиды по прошествии некоторого времени резко начинают вымирать и исчезают из популяции. Почему же?
Оказывается проблема кроется в накоплении непроявляющихся в фенотипе мутаций. Ну например есть у тебя две хромосомы. В одной у тебя рабочий ген А, а вот во второй - уже мутировавший и нерабочий его родственник, ген а. То есть ты гетерозиготный по этому гену и пишешься как "Аа". И вот настало время твоего бинарного деления. Обе твои хромосомы удваиваются и по две расходятся в две дочерние клетки. Варианты полученных клеток могут быть такие (АА + аа) и (Аа+Аа). Так как клетка с генотипом "аа" не будет содержать рабочего гена А ни в одной из хромосом, то она окажется нежизнеспособна и постепенно просто погибнет. Так начнется вырождение полиплоидной популяции.
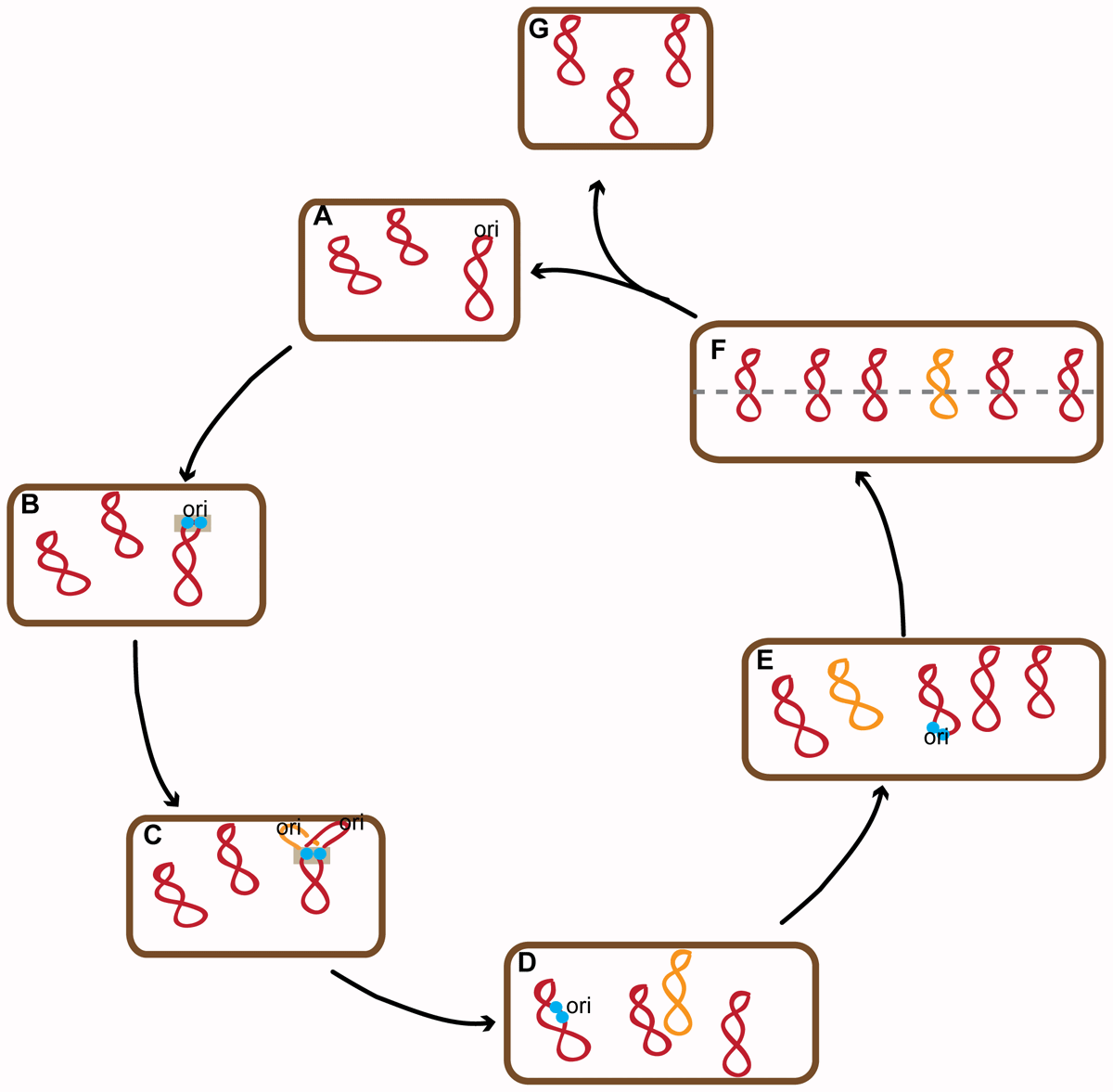
Еще более интересный пример приводит Александр Марков в своей статье, где он предлагает модель показывающую эволюционную судьбу популяций с различной плоидностью при высокой скорости мутагенеза.
У нас есть триплоидная бактерия, её генотип написан слева в столбик. Ну и собралась она значит делиться. Как вы видите в среднем столбике, после репликации мы получаем удвоенный набор хромосом, которые затем в случайном порядке, но по три штуки разбегаются по расходящимся двум клеткам. И вот, после окончания деления оказывается, что одна из бактерий не имеет рабочего гена С, в то время как вторая не имеет рабочего гена А - очевидно, что долго они не проживут и станут так называемым сегрегационным мусором.
Выходит парадоксальная ситуация. Сначала полиплоиды резко распространяются захватывая пищевые ресурсы и вытесняя моноплоидов, а затем, накопив кучу мутаций вымирают сами, что приводит к вымиранию всей популяции. Эволюция ведь вроде работает не так? Или все-таки так? И что же с этим делать? А выход тут один - нужен МИТОЗ.
Но митоз - это ведь сложно, его нельзя изобрести по щелчку пальца, скажет любой из вас. Ведь должны же быть и другие способы! Ну и да, бактерии бы не были самыми распространенными существами на планете, если бы их эволюционный путь закончился бесконтрольными мутациями.
А ЧТО КРОМЕ МИТОЗА?
И первый способ - это выработка циклов плоидности. Здесь всё довольно просто - достаточно просто чаще делиться, иногда пропуская стадию репликации хромосом. Это будет приводить к выявлению мутантных моноплоидов, нерабочие гены которых будут вымываться из популяции. Такие вот американские горки. Стал полиплоидом, захватил зону обитания, потом начал резко делиться, стал моноплоидом, почистил генофонд, затем опять в полиплоида и вперед дальше захватывать мир. НЕИМОВЕРНО УВЛЕКАТЕЛЬНО,
Второй способ - рекомбинация гомологичных участков хромосом, которую можно осуществлять к примеру методом генной конверсии, когда рабочая копия гена из одной хромосомы заменяет и затирает нерабочую версию из соседней. Если генная конверсия будет происходить достаточно активно и при этом явно будет обгонять темпы любого мутагенеза, то полиплоидной популяции практически ничего не грозит. Используется такой метод у многих бактериальных организмов - от метаногенных бактерий до пластидов растений, которые кстати тоже внезапно полиплоидны.
Третий способ - это комплекс мер связывающих вместе горизонтальный перенос генов и кроссинговер. Горизонтальный перенос - это способность бактерий обмениваться участками ДНК между собой, то есть между клетками, кроссинговер - это когда гомологичные участки хромосом меняются местами, но уже внутри одной клетки. Совместив два этих механизма можно будет спокойно собирать рабочие хромосомы и не париться по поводу накопления мутаций. В конце концов всегда можно использовать и генную конверсию, не так ли?
Правда с горизонтальным переносом генов есть один важный косяк - проблема в его асимметричности. Тут дело в том, что когда какой либо ген переходит в другую клетку, то там он затирает свой аналог, вставая на его место. Таким образом, может вполне себе случиться ситуация, когда не очень то и полезный ген получит карт-бланш на распространение, даже в том случае, если не будет особо полезен. Тот же Марков приводит в пример два аллеля - один из них способствует горизонтальному переносу, а второй препятствует. Кто же победит в отборе? На первый взгляд, сам по себе горизонтальный перенос полезен, а значит должен победить способствующий ему аллель. Но на деле, в рамках моделирования, было показано, что ген препятствующий переносу просто со временем перетрет конкурирующий полезный аллель. Даже при учете его, я как я уже сказал, очевидной полезности для организма. Как обойти такое эволюционное недоразумение? Ну например обмениваться целиковыми хромосомами, а обмен участками оставить на откуп кроссинговеру.
С кроссинговером тоже косяк - кольцевая ДНК не очень то хорошо подходит для перетасовки гомологичных участков, да и место начала репликации там чаще всего одно. Точнее сказать сама то кольцевая хромосома для всего почти что подходит прекрасно, но вот только не для кроссинговера. А значит нужны линейные хромосомы однако, да еще и кучей точек репликации!
Ну и четвертый способ - это тот, от которого не отвертишься, нужен митоз. Именно умение не рандомно распихать реплицированые хромосомки по дочерним полиплоидам, а детально и точно скопировать и ровненько и предсказуемо распределить решает проблемы связанные с накопление мутаций и упомянутым выше сегрегационным грузом. То есть при делении дочерняя клетка должна получать все копии генов своего родителя, а не случайный набор реплицированных хромосом.
Но митоз не был бы таким крутым решением, если бы не имел проблемы, которые пришлось решать чуть ли не на ходу. И решением этих проблем стало появление мейоза и полового процесса.
БЕЗ МЕЙОЗА НЕТ РАЗНООБРАЗИЯ!
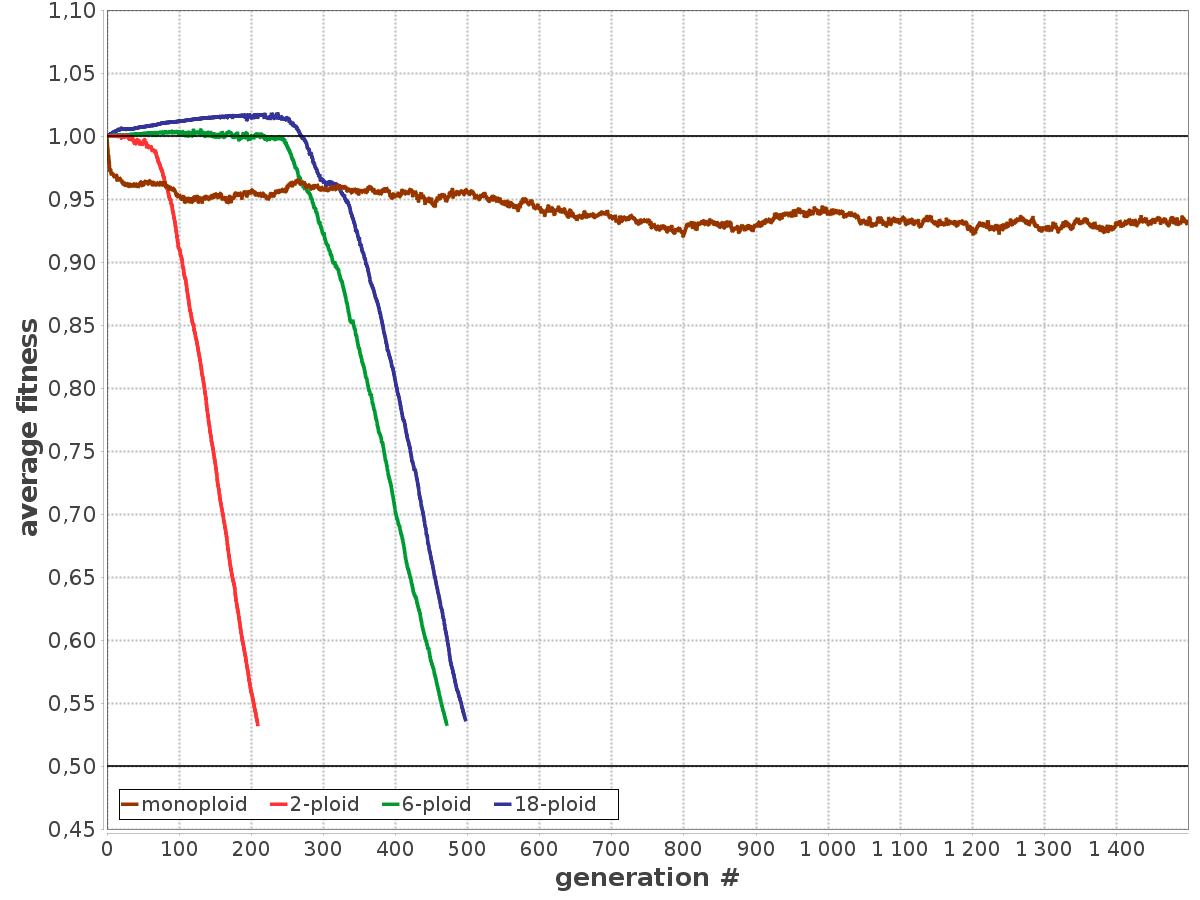
Короче говоря, когда археи таки научились правильно попарно растаскивать реплицированные хромосомы по дочерним клеткам и таки смогли сделать что-то похожее на митоз возникла маленькая проблема - теперь у тебя куча точно скопированных генов и БОЛЬШИНСТВО ИЗ НИХ ЛИШНИЕ. И правда, теперь избыточные копии генов, которые остались от полиплоидной молодости получают необычайную свободу - мутируй как угодно, ведь в какой-то хромосоме есть правильная копия, доставшаяся от родаков и всё считывается с нее. Из этого вытекает один интересный момент - теперь каждая хромосома уже становится банально незаменимой, её нельзя терять, а то ведь вдруг там находится нужный ген, которого больше нигде не сохранилось? Самое интересное, что это не какие-то умозрительные заключения. Абсолютно нет - это показывают результаты моделирования популяции шестиплоидных микроорганизмов. А результаты вот они:

Моделирование проводилось на протяжении тысячи поколений. В каждом поколении гены мутировали, а "качество" этих генов выражается в числе от 0 до 1. Зеленым отмечены самые качественные и маломутировавшие гены. Из этой таблицы видно, что по итогу в каждой хромосоме формируется свой набор удачных генов, а значит для нормальной работы микроба нужны все шесть хромосом - уже не отвертеться.
Это кстати отвечает сразу на два вопроса. Во-первых у предковых эукариот предполагалось наличие множества паралогов (генов-копий), а во-вторых, такое количество "лишней ДНК" объясняет общую тяжеловесность генома эукариот по сравнению с обычными прокариотами. Выходит, что с появлением митоза бывшая полиплоидная архея превращается в моноплоидного протоэукариота с кучей генов-паралогов. Ну а как вы прекрасно понимаете, гены умеют менять места своей "работы", что только лишний раз позволило усложнить строение Последнего Общего Эукариотического Предка (LECA).
Так как теперь у вашего протоэукариота каждая хромосома индивидуальна и содержит свой набор генов, то выходит, что классический горизонтальный обмен с перетасовкой одной двумя случайными хромосомами уже не работает, так и загнуться можно. Значит, нужен более контролируемый способ! И самый простой вариант - это мейоз.
На самом деле мудрить с половым процессом протоэукариотам не требовалось - достаточно осуществить сингамию, то есть слияние двух клеток. После слияния полученная клетка становится диплоидной, осуществляет рекомбинацию хромосом, кроссинговер между гомологичными участками для лучшего перемешивания генного материала, а затем расходится обратно на две клетки. Вот такое вот поддержание генетического разнообразия. Прошли мы с вами путь от обычной бактериальной конъюгации к практически стандартному мейозу, как у нормальных людей. Так что же в итоге?
А по сути получается, что с помощью относительно несложного моделирования и компиляции уже имеющихся данных удается воссоздать эволюцию генетического аппарата от простецкой бактерии к настоящим эукариотам и при чем очень и очень непротиворечиво.
КОРОЧЕ, СКЛИФОСОВСКИЙ!
Сначала у нас обычные полиплоидные археи без митоза. Живут они в тяжелые времена первой кислородной "катастрофы", в довольно экстремальных условиях, где наличие запасных генов полезно из-за постоянных повреждений ДНК. Но чтобы полиплоиды не вымерли из-за мутаций, древние археи вырабатывают механизмы обхода данной проблемы - моноплоидные поколения, унификацию хромосом методом генной конверсии, а также активно работают над постоянным горизонтальным переносом, может даже и через слияние клеток. Затем, чтобы решить проблемы с мутагенезом окончательно, обмен начинает происходить целиковыми хромосомами, а вместо всяческих передач плазмид и кусочков хромосом начинает активно использоваться кроссинговер, что в дальнейшем приводит к появлению линейных хромосом. Далее был запущен процесс митоза, который решал проблему сегрегационного груза за счет попарного разделения реплицированных хромосом между дочерними клетками. Закрепление митоза в популяциях привело к тому, что постепенно происходит диверсификация и специализация хромосом, каждая из них начинает содержать необходимый и незаменимый набор генов, а полиплоидный организм постепенно становится моноплоидным. Это приводит к тому, что стандартный горизонтальный перенос перестает работать и для полового процесса начинает использоваться рекомбинация между гомологичными хромосомами среди ближайших по родству клеток. Постепенно выделяются отдельные популяции, хромосомы которых уже не способны рекомбинировать, что приводит к развитию репродуктивной изоляции и появлению ПЕРВЫХ НА ПЛАНЕТЕ БИОЛОГИЧЕСКИХ ВИДОВ.
Вот так мы и прошли путь от обычно моноплоидной бактерии до предка эукариот. Ну а далее вы уже знаете - симбиотическая теория.... ИЛИ НЕ ЗНАЕТЕ? Если не знаете, то предлагаю перейти в комментарии и сказать, нужно ли продолжение! =)
СТРИМЫ: https://trovo.live/crazypaleo
ПОДДЕРЖИ НА БУСТИ: https://boosty.to/crazypaleo