Когда речь заходит об эволюции, сторонники креационизма часто любят говорить о том, что "переходные формы не найдены". В действительности, переходных форм организмов, иллюстрирующих постепенные изменения биосферы, довольно-таки много. Сегодня мы расскажем об одной из самых очевидных эволюционных цепочек. Очевидных - в том смысле, что довольно сложно как-то ещё объяснить такую последовательность существования живых организмов, постепенно накапливающих определённые признаки. В нашем случае - это эволюция китов. Итак, поехали!
Откуда пошли китообразные

Традиционные взгляды на эволюцию китообразных состояли в том, что их ближайшими родственниками и, вероятно, предками являлись мезонихии - вымерший отряд хищников, которые напоминали волков с копытцами вместо когтей и были близкими родственниками привычных нам парнокопытных зверей - коров, коз, свиней и других. Эти животные имели зубы необычной конической формы, похожие на зубы китообразных. В частности из-за этого учёные долгое время считали, что китообразные произошли от некой предковой мезонихии.
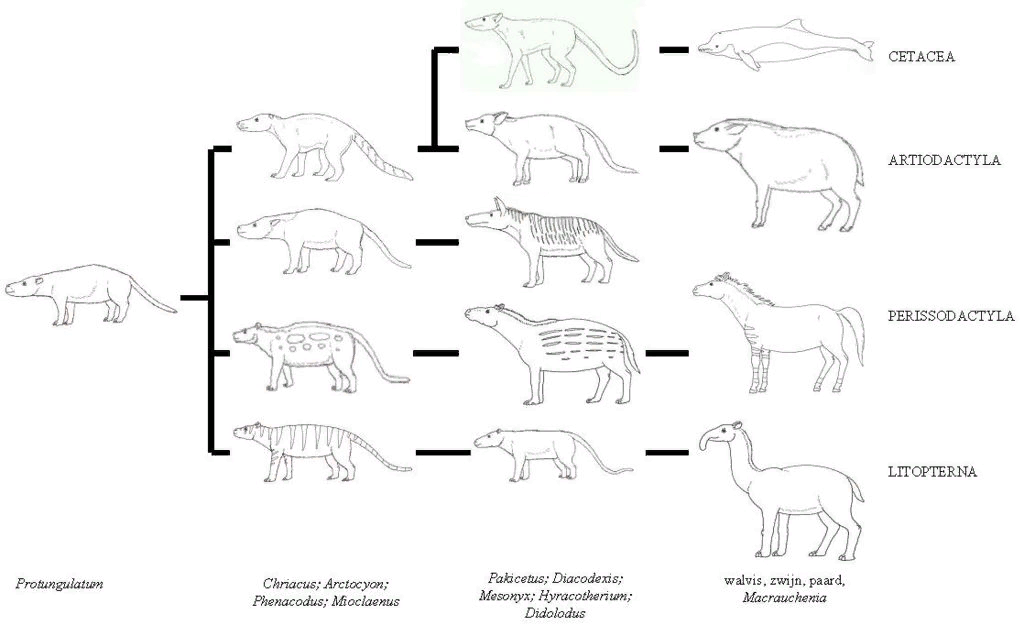
В 1985 году Винсент Шарич провёл молекулярно-генетическое исследование белков крови, результаты которого свидетельствовали, что китообразные — близкие родственники парнокопытных, в частности гиппопотамов. В 2005 году палеонтолог Жан-Рено Буассери и его коллеги Мишель Брюне и Фабрис Лихоро провели анализ новых и ранее существовавших окаменелостей бегемотов, китов и антракотериевых и убедительно доказали их близкое родство.
Итак, какие же переходные формы существовали на пути китов с суши в океаны?
Пакицет
Пакицеты были копытными животными, иногда их классифицируют как ранних китов. Они жили на территории современного Пакистана (откуда и название — «кит из Пакистана») в раннем эоцене, около 50 миллионов лет назад. Это было животное, внешне напоминавшее собаку, однако с копытами на пальцах и с длинным тонким хвостом. С китами пакицета роднит устройство уха: слуховая булла пакицета, как и у кита, образовалась исключительно из барабанной кости. Форма ушной области пакицета весьма необычна и находит аналоги только у китообразных. Первоначально предполагалось, что ухо было приспособлено для жизни под водой, однако, как показали дальнейшие исследования, уши пакицета подходят только для воздушной среды, и, если пакицет действительно предок китов, возможность слышать под водой была позднейшей адаптацией уже имевшегося слухового аппарата. Как утверждает Тевиссен, зубы пакицета тоже напоминают зубы ископаемых китов.
Индохиус
Учёные обнаружили, что устройство уха, как у пакицета, наблюдалось в окаменелостях небольшого похожего на оленя животного индохиуса (правильно, на самом деле, говорить "индохий", но в русскоязычной литературе укоренился вариант с суффиксом "ус"). Индохиус обитал около 48 миллионов лет назад в Кашмире. Это небольшое — размером с домашнюю кошку — травоядное животное обладало некоторыми чертами, сближающими его с китами и свидетельствующими об адаптации к водной среде. В их числе толстая и тяжёлая костная оболочка, напоминающая костную оболочку некоторых современных полуводных животных, таких как гиппопотамы, что способствует уменьшению плавучести и, как следствие, позволяет оставаться под водой. Это позволяет предположить, что индохиус, подобно современному водяному оленьку, нырял под воду, чтобы укрыться от хищника.
Амбулоцет
Наиболее примечательным из древних китов является амбулоцет, известный из эоцена Пакистана. Внешне это млекопитающее походило на трёхметрового крокодила. Амбулоцет был полуводным животным: его задние лапы лучше приспособлены для плавания, чем для передвижения по суше. Вероятно, он плавал, изгибая тело в вертикальной плоскости, как современные выдры, тюлени и киты. Предполагается, что амбулоцетиды охотились подобно современным крокодилам, подстерегая в засаде рыб и пришедших на водопой животных. Амбулоцет отличался от пакицета и индохиуса тем, что был более приспособлен к водному образу жизни: скорее всего, большую часть времени он проводил в воде, имел развитые перепонки и короткие лапы, начавшие преобразовываться в ласты. В то время как пакицет был больше сухопутным животным, которое хоть и хорошо плавало, но основную часть жизни ходило по суше.
Ремингтоноцетиды
Более мелкими родичами амбулоцетид были ремингтоноцетиды, известные из Индии и Пакистана, из отложений возрастом 49–43 млн лет назад. Эти животные обладали сильно вытянутыми челюстями и были несколько лучше приспособлены к подводному плаванию, чем амбулоцетиды. Они тоже имели хорошо развитые конечности, но отличались маленькими глазами, тонкими челюстями и расширенным основанием черепа. Если сравнивать «земноводных китов» по общему облику с крокодилами, то среди них амбулоцетиды больше напоминали аллигаторов, а ремингтоноцетиды - гавиалов. Строение слуховой области в сочетании с маленькими глазами показывает, что для обнаружения добычи ремингтоноцетиды пользовались в основном слухом. Известно шесть родов ремингтоноцетид. Их размеры были невелики: например, качицет не превосходил величиной речную выдру. Для эндрюсифия установлено наличие мощного уплощенного хвоста, служившего основным органом движения в водной среде. Райанист (Rayanistes afer), найденный в среднем эоцене Египта, демонстрирует, что ремингтоноцетиды достигли южного побережья Тетиса.
Протоцетиды
Первыми с мелководья на настоящую глубину отправились протоцетиды, жившие 49–37 млн лет назад. Их главным эволюционным приобретением был удлиненный мощный хвост (поначалу без хвостовой лопасти), который обеспечивал быстрое плавание. При этом протоцетиды сохраняли развитые задние конечности. Вероятнее всего, по образу жизни их можно сопоставить с современными крупными ластоногими. У некоторых протоцетид зубы были уменьшены, это свидетельствует, что добычу - мелких морских животных — они заглатывали целиком. Такими были первые китообразные, которые сумели распространиться за пределы моря Тетис - их остатки найдены в мелководно-морских отложениях в Африке, Европе и Северной Америке. Самые крупные из них достигали длины более 3 м и весили до 400–500 кг. Вероятно, от протоцетид произошли все более поздние китообразные, включая современные группы. Описано около 20 родов протоцетид, из них наиболее известны протоцет (Protocetus) и родоцет (Rodhocetus). Своеобразно выглядел макарацет (Makaracetus bidens), у которого (судя по строению передней части черепа) был короткий хоботок или мясистая верхняя губа, которые могли служить для сбора со дна моллюсков и других животных.
Базилозавриды и дорудонтиды
Базилозавриды и дорудонтины жили вместе в позднем эоцене примерно от 41 до 33,9 миллиона лет назад и являются древнейшими известными облигатными водными китообразными. Это были полностью узнаваемые киты, которые физически не могли долго передвигаться по суше. Это подтверждается их окаменелостями, обычно обнаруживаемыми в отложениях, свидетельствующих о полностью морской среде, в которой отсутствует приток пресной воды. Они, вероятно, были распространены по всему тропическому и субтропическому морям мира. Базилозавриды обычно встречаются в ассоциации с дорудонтинами и были тесно связаны друг с другом. Окаменелое содержимое желудка одного базилозаврида указывает на то, что он ел рыбу.
Хотя они очень похожи на современных китообразных, базилозавридам не хватало "дыни" - округлого выступа на морде, который позволяет зубастым китам использовать эхолокацию. У них был маленький мозг; это говорит о том, что они были одиночками и не имели сложных социальных структур некоторых современных китообразных. Их орбиты были обращены вбок, а носовое отверстие переместилось еще выше по морде, ближе к положению дыхала у современных китообразных. Кроме того, их ушные структуры были функционально современными, со вставкой заполненных воздухом пазух между ухом и черепом. В отличие от современных китообразных, базилозавриды сохранили большой наружный слуховой проход.
У обоих групп есть скелеты, в которых сразу можно узнать китообразных. Базилозавриды были размером с более крупных современных китов, например базилозавр достигал длины до 60 футов (18 м) в длину; дорудонтины были меньше, типовой род - дорудон - достигал примерно 15 футов (4,6 м) в длину. Большие размеры базилозаврид обусловлены крайним удлинением их поясничных позвонков.Передние конечности базилозаврид, вероятно, имели форму плавников, а задние конечности были крошечными и, безусловно, не участвовали в передвижении. Однако они сохранили подвижные суставы своих родственников, которые могли передвигаться по суше. Две крошечные, но хорошо сформированные задние лапы базилозаврид, вероятно, использовались в качестве класперов при спаривании. Тазовые кости, связанные с этими задними конечностями, не были соединены с позвоночным столбом, как это было у протоцетид. По существу, любые крестцовые позвонки больше нельзя четко отличить от других позвонков.
К современным китам
Первые признаки, указывающие на возможную эхолокацию, были обнаружены у олигоценового зубатого кита котилокары из вымершего семейства ксенорофид, он жил у побережья Южной Каролины где-то 28 млн лет назад. В ту эпоху было очень много разных китов, имевших примитивные признаки - зубы, похожие на таковые у полусухопутных предков китов, но не у современных китообразных. У эвринодельфиса из семейства эвринодельфинид беззубый конец верхней челюсти превратился в длинный отросток, служивший смертоносным оружием, как у рыбы-меча. Этот «мечерылый дельфин» был небольшим (около 2 м), но родственный ему макродельфин достигал размеров косатки (6–8 м). Сквалодон из семейства акулозубых дельфинов имел длинные узкие челюсти. Позднее начали появляться виды, которые внешним видом весьма напоминали современных дельфинов, появившихся около 15 млн лет назад.
Появление дельфинов
Симоцет из верхнего олигоцена Орегона (США), выделенный в самостоятельное семейство симоцетид, получил прозвание «курносый дельфин». Его короткие челюсти были заметно загнуты вниз, а редко расположенные зубы служили скорее для просеивания пищи, чем для ее разрывания. Вероятно, симоцет добывал донных беспозвоночных, процеживая осадок.
У двухметровой морской свиньи семиростра (Semirostrum ceruttii), жившей 3 млн лет назад у берегов южной Калифорнии (США), нижняя челюсть была вдвое длиннее верхней. Эта часть челюсти формировала покрытый кожей выступ-подбородок, который использовался как специальный чувствительный орган для поиска пищи «на ощупь» в мутной воде у самого дна .
История группы речных дельфинов (Platanistoidea) прослеживается с позднего олигоцена, когда ее представители еще обитали в морских бассейнах. При этом оказывается, что они вовсе не близкородственны друг другу. Слепой гангский дельфин (Platanista) принадлежит линии архаичных зубатых китов, родственной сквалодонтидам, тогда как другие - амазонский, лаплатский и озёрныйLip — скорее всего, происходят от разных вымерших групп.
Кашалоты
Ныне это специализированные охотники на кальмаров. Однако некоторые неогеновые кашалоты были китами-убийцами своего времени, занимая экологическую нишу косаток до их появления около 3 млн лет назад. "Кашалотом-убийцей" часто называют зигофисетера из позднего миоцена Италии, возрастом 10–7 млн лет. Сие чудище было длиной 6–7 м, имело мощный череп и многочисленные большие зубы. Были и иные грозные: семиметровый бригмофисетер из Японии (15–14 млн лет) и представители рода акрофисетеров из Перу, достигавшие в длину лишь 4 м, но также занимавшие нишу активных морских охотников.
В Перу были обнаружены останки огромного морского хищника. В длину гигантский кашалот-убийца достигал 18 м, а весил он около 40 т (примерно как самцы современного кашалота). Длина его черепа около 3 м, а зубы имеют диаметр около 12 см и длину до 36 см. Пожалуй, это самые крупные зубы в животном мире, не считая гипертрофированных клыков и бивней. Колосс получил название "мелвиллов Левиафан" (Livyatan melvillei), в честь морского монстра левиафана из Библии и американского писателя Германа Мелвилла, автора романа «Моби Дик, или Белый кит». Можно предполагать, что появление таких крупных хищников было связано с развитием гигантизма среди их потенциальных жертв — усатых китов.
Усатые киты
Примерно 30 миллионов лет назад, в олигоценовую эпоху, появились киты, которые мирно плавают в океане и фильтруют планктон с помощью своего китового уса. Интересно, что у китов группы эомистицетид (Eomysticetus, Waharoa и др.), сравнительно некрупных (до 8 м) животных стройного сложения, китовый ус занимал только заднюю часть неба, на переднем конце челюсти имелось несколько зубов, а дыхало располагалось на значительном расстоянии спереди от глаз, что выглядит довольо архаично по сравнению с современными китами. Самые древние гладкие киты появились 23 млн лет назад. Полосатиковые (Balaenopteridae), к которым относится гигантский синий кит, появились в Северной Атлантике около 12 млн лет назад. Судя по ископаемым остаткам, современному синему киту по размерам не уступали некоторые киты-полосатики плиоцена и плейстоцена, например Balaenoptera sibbaldina из раннего плиоцена Бельгии. Однако самыми широко распространенными усатыми китами неогена были цетотерии (Cetotheriidae) — очень разнообразная (более 50 видов и около 20 родов) группа сравнительно небольших форм (около 3–4 м) без признаков специализации современных семейств.
А на этом всё! Подписывайтесь на канал, ставьте лайки, делитесь публикацией в соцсетях, не забывайте заходить на вики и к нашему другу - Упоротому Палеонтологу! Особая благодарность тем, кто пожертвовал нам на развитие проекта, мы это очень ценим!
Также предлагаем вам оценить ещё один канал, созданный вторым редактором Вымершие животные вики, посвящённый различным мифам и легендам, древним и современным.
#Киты #интересные факты #эволюция #наука #биология #палеонтология #просто о сложном