Зарождение Жизни
Сколько громких заголовков о зарождении Жизни (ЗЖ) мы видим последние годы. Находят органические молекулы и то, достаточно сложные, аминокислоты и т.п., на астероидах в газовых и пылевых звёздных облаках. Панспермия (появление Жизни на Земле из Космоса) рассматривается как ведущая гипотеза в.. СМИ. Ученые редко вступают в споры в СМИ. Спорить тут надо с цифрами на руках. Наука сегодня находится на достаточно высоком уровне, чтобы доказательно ответить на этот вопрос. Среди биологов-генетиков основная модель сегодня: RNA (рибонуклеиновые кислоты) модель Зарождения Жизни.
Существующие модели появления жизни на Земле должны ответить на главные вопросы:
1-Движущие силы - `автопилоты` до появления клетки?
2- Вероятность возникновения и путь? ( Земля | да и Вселенная | слишком молода , чтобы обеспечить вероятность возникновения жизни из белков).
3.-Предложить вектор развития Жизни.
КАК же это случилось? Описываемые в работе химические реакции, конечно же, являются одними из огромного числа, но выделяются, как наиболее вероятные и важные для описания процессов приведших к ЗЖ на Земле. Основная идея, очевидность ЗЖ, через информационную часть. Т.е., `цель` жизни и ее суть - самовоспроизведение, именно с него и началась история жизни на Земле.
3,9 млрд. лет до НЭ.
Земля остывает после гравитационного сжатия, избыток восстановителей превращает её в хороший химический реактор. Главные восстановители – гидриды металлов, металлы из-за своей плотности ушли в глубину Земли. Легкие элементы и их соединения оказались выжаты в верхние слои планеты. Таким образом, создалась возможность для сохранения кисло-рода в газообразных соединениях типа СО, СО2, НСОН и у поверхности планеты обеспечивалась высокая концентрация углерода и его производных. Все эти возможности с легкостью реализовывались в лабораторных работах при моделировании условий на Земле времен 4 млрд. лет назад. Соответственно обычными производными были нитрилы и аминопроизводные, молнии и горячие процессы около большого количества вулканов в присутствии переходных металлов – были мощными реакторами. Сахара тоже легко производились в такой системе. Именно поэтому поверхность древнейшей Земли была в избытке наполнена углеводородами. Их происхождение было абиогенным, карбонаты восстанавливались водородом при высоких давлениях и температурах. Также в самом начале гидриды металлов взаимодействовали с оксидами углерода, в присутствии воды, что также давало углеводороды и другие сложные органические производные. Также образовывались, карбиды, которые опять же разлагались водой с образованием углеводородов.
Чем RNA (РНК - рибонуклеиновые кислоты) живее другой органики?
Дело в том, что уже у достаточно короткие полимеры РНК (а РНК и ДНК – это полимеры) обнаружили удивительные катализаторные свойства. Они могут помогать протекать органическим реакциям, помогать расщеплять себе подобных и наоборот – синтезировать себе подобных и самих себя. Они участвуют в сплайстинге, конструировании меседжРНК, которые поступят в рибосомы и напечатают записанную в них информацию, превратив её в белки. Более того - они могут уже конкурировать между собой за пищу! Их пища нуклеотиды – мономеры, кусочки из чего состоят они сами. Более того, они способны на эволюцию, ну чем не кандидаты в первые «разумные» молекулы?
Тут следует сказать, что ученые не сразу узнали об этих способностях микроРНК. Уже будучи в числе первых кандидатов в родоначальники Жизни они постепенно обросли многими удивительными подробностями своих способностей, что только укрепляло их позиции. Ведь что самое главное в поиске НАЧАЛА? Найти простейшую комбинацию из простейших компонентов позволяющих запустить старт Жизни. Тут с РНК невозможно поспорить никаким другим классам веществ. Только четыре (!) молекулы могут оживить органику. При этом две из них простейшие: Фосфорная кислота (или фосфаты) Рис 1, гликоль рибоза Рис 2.
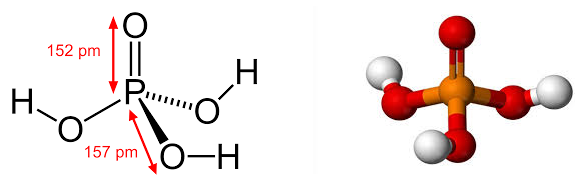
Рис 1 Фосфорная кислота
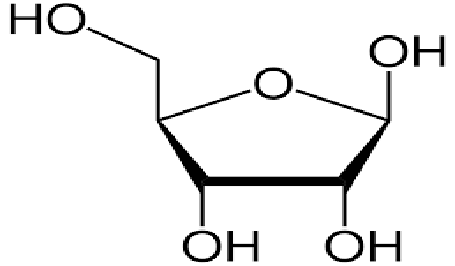
Рис 2 Гликоль молекула Рибозы
и два нуклеиновых основания (НО) Рис 3 и Рис 4 :
Рис 3 Аденин
Рис 4 Урацил
Более простой комбинации придумать просто невозможно. Три органических химиката и фосфорная кислота в растворах и реактор жизни – запущен.
Вся проблема в первичном «оформлении» старта. Требования к «ассортименту» минимальные, всего четыре типа простых молекул и органика - оживает.
Теперь попробуем ответить на часть вопроса по движущим силам:
Конечно же, недопустимо вводить биоорганические (живые) атрибуты для объяснения входа в химию Живой органики. Для нашего случая этого не требуется, все четыре действующих лица – достаточно простые вещества. Синтезирование их природой имеет только проблемы с выходом (урожаем) производных и то только двух нуклеиновых оснований (НО). Но природные «концентраторы» - туфы, силикаты, глинозём и фосфаты - в неисчислимых количествах присутствуют на Земле. И вот уже на втором шаге после синтеза нас ждут эти особые свойства микроРНК, биорганика оказалась вплетённой в химию НО.
Так, основным вопросом становится уже разбор этих трёх первых шагов по элементам.
1 – синтез нуклеиновых оснований (аденин, урацил),
2 – синтез нуклеотидов – мономеров РНК (ДНК).
Если первый вопрос упирается только в выход продукта в химических реакциях, то второй имеет и стереоскопические аспекты. Так, НО могли сорбироваться на фосфатах и силикатах.
Пример :
Рис 6
Вот так (Рис 6) выглядит система внутренних связей Нуклеиновых Оснований (НО), которая и обеспечивает на уровне минимолекулярных образований биологические свойства Живых молекул. Заметьте, это происходит уже в самом начале усложнения такой системы. Для простоты, представим себе нечто похожее, но вместо известных нам Аденина и Урацила будут похожие пронуклеиновые основания. А вместо фосфорной кислоты – матрица из кремнезема или фосфатов, матрица временно решает вопрос и с гликолями. Пронуклеиновые основания могли прямо сорбироваться на матрице. Против них выстраивались Комплементарные пары, и вот уже в таком положении они могли пространственно организовать удобную для сшивки компоновку. Рис 7
Рис 7
Очевидно, что RNA были основой возникновения клеточной структуры. Как и последующая эволюция: эволюция матрицы - DNA и RNA молекул. Как будет показано ниже, время возникновения первых про - RNA молекул, было исторически коротким. Вероятность возникновения жизни настолько высока, что придется принять, что Жизнь, в ее простейшем виде - самовоспроизведение по этой модели, может возникнуть всюду где условия даже периодически повторяют `активные циклы` ( т.е. если даже основные температуры низкие, но существуют периоды с температурами 10С < Т < 100С, с присутствием нижеперечисленных компонентов в атмосфере и на поверхности , и с ограниченными зонами перегрева ). В дальнейшем изложении своей теории, автор рассматривает только значимые для возникновения жизни процессы и реакции. В данном случае рассматриваются системы Земного типа.
----Можно классифицировать и эволюционировать историю развития Жизни на Земле:
Используемые сокращения :
DNA- дезоксирибонуклеиновая кислота;
RNA- рибонуклеиновая кислота;
PUO(ПУО) - Пуриновые основания;
PMO(ПМО)- Пиримидиновые основания;
p-Nzd - пронуклeозиды;
p-Ntd - пронуклeотиды;
pNtd - полинуклeотиды;
Gl гликоли;
PAR про- реактор;
IE , ИЕ информационные единицы.
A-IB - блоки pNtd не содержащие информации.
А. Накопление простейшего органического материала (углеводородов).
Б. Накопление сложных органических веществ в том числе гетеросоеди-нений. Появление ПУО и ПМО и их концентрирование на силикатах и полифосфатах ;
С. Накопление p-Nzd и p-Ntd и их концентрирование. Адсорбция их на силикатах и полифосфатах ;
Д. Транскрипция поли-пронуклеотидов на них ;
Е. Увеличение степени полимеризации ,появление ИЕ;
--Примерный расчет:
Можно оценить поверхность про - реактора ( PRO-ALIVE Reactor- `PAR`) - который образовывался из уже известных нам пород например : микро-пористого силикогеля ~ 10 кв.м./ ( гр.минерала ) или -10 degr(7) кв.м./ (тон.минерала). Выбор минерала основан на близости структурных постоянных связей силиката к существующим комплементарным связям - 3,4 А / в действительности, с учетом `экзотических `гликолей ,`работали` и другие размеры / Рис 5.
При величине природного минерального образования (малого размера) ~ до 100 тон ( около 7х6х7 метров) ( активной плотностью до 0,3), можно оценить поверхность одного PAR как -10 degr(9) кв.м.. При постоянной (-Si--O--Si-) , I ~ 3,4 А - получим плотность активных (РИС.3) центров ~ 3*10 degr (17)/кв.м.. Или для одного реактора, ~3*10degr(28) активных центров в виде ковров. Для заполнения одного такого- PAR необходимо ~ 3*10degr (28) молей нуклеиновых оснований (при заполнении 10% -3*10degr(27) молей , или около 2х тон смеси ( PMO и PUO ).
Такой реактор служит `фабрикой`, где идет самодоработка полинуклеотидов. Стерическое автопилотирование реакций полимеризации, так можно назвать эти процессы. Учитывая селективность процессов, необходимо признать, что предложенный механизм укладывается в требования логики и теории. Вероятность появления определенной последовательности DNA из 40 нуклеотидов ~ 4 degr(40) или ~ 10 degr(24) - пере-боров. Что может быть легко обеспечено уже при заполнении PAR. Можно оценить производительность обменов типа:
Si-O--PMO + PUO ---> Si-O--PUO + PMO
РИС. 8
Примем скорость обмена ~ 1 обмен/сек. Для нуклеотида из 60 оснований - требуется вероятность ~ 4 degr(60) ~ 10 degr(36) , что обеспечится таким PAR за ~10 degr(9) сек или ~ 30 лет.
Ясно, что остановки случайны, а обмены происходят циклически -волнами . Период волны ~ геологическим эффектам, например: волны от водоема (гейзеры и т.п.) + дожди. Получим min ~10 сек, max ~ 1 час. Средний пери-од ~ 30 мин. Т.о., в вышеописанном случае / скорость генерации для 60 оснований / ~ 30 лет х 1800 ~ 0,054 млн.лет .
Но учитывая поливариантность, видимо, этот срок ~ 100 раз меньше. Учитывая также низкий выход реакций / 3` - 5`/ ; и др. побочные процессы , конкурирующие реакции – уменьшение на 2 порядка, не селективность фосфатирования – уменьшение на 2 порядка , др. факторы 2 порядка / эта цифра увеличится ~ на 5 порядков, т.о., будем иметь ~ 50 млн. лет. Ясно, что такие системы будут `перегружены` бессмысленной информацией. Эти огромные довески, можно грубо оценить на одну IU ( про-ген ), как равному порядку поливариантности, т.е., приблизительно ~ 2-4 порядка. Для грубой оценки , можно взять величину гена нуклеазы 600 пар Ntd для него A-IB достигает от 60 000 пар до 6 млн. пар Ntd.
Ж. Появление `композитов`- шли естественно-случайные сборки липидов , нуклеотидов, полинуклеотидов, металлокомплексов, аминокислот и других органических соединений в `композиты`- которые начинают участвовать в процессах обмена в этой `полинуклеотидной каше.
З. Появление `свободных` полимерных молекул полипродезоксинуклеотидов и поли-прорибонуклеидов накопивших необходимые ИЕ обеспечивающие цикл самовоспроизводства и "кооперация" с другими дополняющими ИЕ. Естественные оболочки - микропузырьки в минералах, были первыми `клетками, в которых и обмен с окружающей средой иногда становился специфичным - по микротрещинам в ней. Пузырьки позволяли практически идеально `доваривать` смеси полинуклеотидов и органики попавшие в них. Можно представить, что такая микротрещина играла роль селективной мембраны. Т.к. ее раз-меры и состав селективно определяли скорость движения различных типов молекул. Не удивительно, что первые системы были `привязаны` к силикатам - еще и потому, что они легко и различны-ми путями образуют пузырьки ( лава, вулканический пепел, перегрев силикатных растворов и т.п.) с большими удельными поверхностями полузакрытого типа. Естественно ожидать, что первыми заработали силикатно-активные органические `композиты` - они сразу изменяли ситуацию в `клетке` - позволяя изменять ее селективные свойства, селективно сорбировались на стенках `пузырьков.
Автор считает, что описываемая выше схема - Твердофазной Матрицы - полностью соответствует требованиям параметров теории и параметров времени, необходимым для описания процессов, приведших к появлению живой материи на Земле. Эта модель дает пищу для понимания процессов эволюции и пути эволюции, в основе которой оказывается эволюция DNA и RNA. Самым неожиданным является вывод о гигантских `довесках` - бессмысленных ( не работающих ) последовательностей, что коренным образом меняет наши представления о пути будущей эволюции и самих механизмов эволюции.
Автор еще раз уточняет, что целью этой работы было: выяснить роль и возможность появления первичной `живой` структуры из автофункционирующего механизма, функционирующего на основе приемлемых начальных условий (с химической точки зрения, геологической, физической и вероятностных критериев процессов , а так же логической достаточности и необходимости).
#Жизнь #зарождение #РНК #происхождение #земля #




















