Появление лекарственной устойчивости обусловлено множеством факторов, в том числе индивидуальными вариациями у пациентов и генетическими различиями в опухолях соматических клеток. Механизмы лекарственной устойчивости и мультирезистентности включают усиленную экспрессию переносчиков АВС, увеличивающих выбросы противораковых препаратов, изменения метаболизма препаратов, мутации лекарственных мишеней и активацию выживаемости или инактивации сигнальных путей смерти ниже по течению. Обычно наблюдается сверхэкспрессия азбучных транспортеров, например, гликопротеина P-170, белка устойчивости к раку молочной железы и белков мультирезистентности в опухолевых клетках после лечения препаратами. Зависимые от АТФ белки транспортера выводят из раковых клеток различные противораковые препараты, в том числе структурно отличные друг от друга.
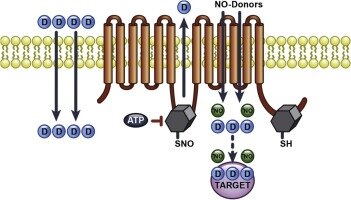
Окись азота, образованная из L-аргинина синтезазой окиси азота, индуцирует гибель различных опухолевых клеток и поэтому, как известно, играет важную роль в лечении онкологических заболеваний. NO - недолговечная молекула свободных радикалов, которая выступает в роли клеточного посланника и играет ключевую роль во многих важных физиологических функциях. Образование реактивных метаболитов, например, NO+, N2O3 и -OONO, было вовлечено во многие действия NO. Эти промежуточные соединения вступают в реакцию с сульфгидрильными группами белков, что приводит к образованию группы S-нитрозотиола - процесса, известного как S-нитрозилирование. Многие опухоли человека характеризуются высокой экспрессией iNOS и выделяют значительное количество NO.
Материалы и методы
1. Клеточная культура
Клетки культивировались в среде RPMI без фенолового красного цвета, дополненной 10% сывороткой крупного рогатого скота и антибиотиками. Камеры обычно использовались для 20-25 проходов, после чего камеры были выброшены.
2. Цитотоксичность и изменение направления исследований резистентности
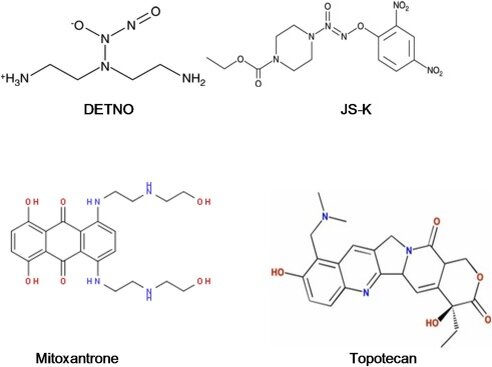
Исследования цитотоксичности проводились с помощью метода ингибирования роста клеток. Вкратце, 100-150 000-150 000 ячеек/лунок были покрыты в 2 мл полной среды на 6-луночный планшет и могли прикрепляться в течение 18 ч. Клетки были трипсинизированы, и количество выживших клеток определялось путем подсчета клеток в счетчике. Для изменения устойчивости клетки сначала обрабатывали JSK или DETNO в течение 2 ч в среде, содержащей 1% FBS без антибиотиков и 200 мкл PBS, затем добавляли различные концентрации препаратов и затем выдерживали в течение 72 ч в среде в целом.
3. Конфокальная микроскопия белка БЦРП в опухолевых клетках MCF-7/MX
Около 1 × 105 родительских WT MCF-7 и устойчивых опухолевых клеток молочной железы MCF-7/MX в течение 18 ч при 37 °C покрывались культурными пластинами на стеклянных покровных пластинах в полной среде. Для исследования наличия или отсутствия белка BCRP клетки фиксировали 4% параформальдегидом в течение 15 мин при комнатной температуре, промывали дважды в течение 5 мин, проникали в течение 5 мин с 0,5% тритоном Х-100 и промывали дважды в течение 5 минут. После блокировки желатином 4% рыбы в PBS в течение 2 ч при комнатной температуре клетки инкубировали с кроличьим анти-BCRP антителом в течение 2 ч, затем вторично антимышечным желатином Alexa Fluor 668 на 1 ч. Крышки с клетками промывали четыре раза и закрепляли на стеклянных горках с помощью антифриза.
4. Молекулярное моделирование
Структуры ABCB1 и ABCG2 рассматривались как рецепторные конфигурации для молекулярной стыковки. Функция скоринга основана на форме лиганда, водородной связи между лигандом и рецептором, взаимодействиях водородной связи с неявным растворителем и взаимодействия металл-хелятор.
Были добавлены протоны, затем добавлены контр-ионы, и полученная структура была растворена в коробке с водой с 50 515 молекулами. До достижения равновесия система состояла из 169 842 атомов, которые были подвергнуты следующим этапам: a) в течение наносекунды ограниченной молекулярной динамики с тяжелыми атомами белка в исходных положениях с постоянной силой 10 ккал/моль/нм, b) минимизация, c) низкотемпературное моделирование MD под постоянным давлением для обеспечения разумной начальной плотности, d) минимизация, e) поэтапный нагрев MD при постоянном объеме и f) постоянный объем MD с шагом уменьшения силы еще на 15 нс.
Цитотоксичность и возврат к лекарственной устойчивости в клетках NCI/ADR-RES по данным JS-K
NO/NO-производные виды ингибируют активность АТФаз и вызывают обратный эффект устойчивости к ADR в P-gp-переэкспрессии опухолевых клеток NCI/ADR-RES. JS-K активируется внутриклеточно системой GSH-GST для генерации NO. В настоящем докладе мы рассмотрели влияние JS-K на реверсирование ADR-сопротивления в NCI/ADR-RES клетках, поскольку NCI/ADR-RES клетки содержат значительно большее количество GSH, чем родительская линия и сверхэкспрессии GST. Несмотря на различия в содержании GSH и экспрессии GST, JS-K одинаково цитотоксичен как для родительского ОВКАР-8, так и для устойчивых NCI/ADR-RES опухолевых клеток.