Начало
Аутофагия, также называемая макроавтофагией, является консервативным деградирующим процессом эукариотических клеток и имеет решающее значение для непрерывного удаления поврежденных белков и эктопических органелл. Особенно в периоды генотоксического стресса, окислительного стресса и голодания питательными веществами аутофагия является классическим клеточным механизмом выживания, который обеспечивает организм необходимой энергией до улучшения состояния. Это также процедура внутриклеточной деградации органеллы и белка, которая включает в себя аутофагосомальное/лизосомальное отделение. Аутофагическая гибель клеток, тип запрограммированной неапоптотической гибели клеток, характеризуется самоеданием безъядерной конденсации, при которой внутриклеточные образования поглощаются в двухмембранных пузырьках, также называемых аутофагосомами, которые затем сливаются с лизосомами для массовой деградации. Двухмембранная структура аутофагосомы может быть получена из эндоплазматического ретикулума, плазменной мембраны, митохондрий, комплекса Гольги или промежуточного отделения ЭРГИК. Комплекс Unc-51-подобной аутофагии, активирующий киназу 1 (УЛК-1), способствует активации аутофагического потока, а комплекс фосфатидилинозитола класса Ⅲ (PtdIns)3-киназы/BECLIN-1 отвечает за зарождение аутофагосомы. Легкая цепочка 3 /аутофагированный белок 8-фосфатидилэтаноламин и комплексы ATG5-ATG12 контролируют удлинение и закрытие аутофагосомных мембран. Удлинение аутофагосом происходит в двух различных клеточных катаболических процессах - вездесущезависимых и вездесущих независимых. На сегодняшний день описаны три типа путей деградации: макроаутофагия, микроаутофагия и аутофагия при посредничестве сопровождающего лица. Функция CMA заключается в доставке дисфункциональных белков и поврежденных органелл, направленных на мотив последовательности KFERQ, в лизосомы путем взаимодействия с лизосомальным мембранным белком Lysosome-ассоциированным мембранным белком 2 (LAMP-2a) и белком теплового шока 70. Таким образом, накопленные данные показывают, что аутофагия играет решающую роль в поддержании клеточного и органического гомеостаза.
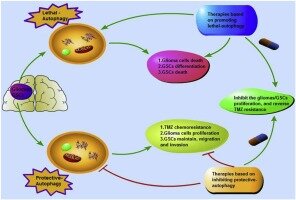
Нейроны представляют собой высокополяризованные клетки с выраженной клеточной конструкцией. Зрелые аксоны являются основным местом для передачи сигнала на другие клетки и содержат обильные ER, лизосомы и эндосомы, однако, есть отсутствие комплекса Гольги, низкая активность деградирующих ферментов и слабее кислотных условиях в зрелых аксонах. Во многих типах клеток аутофагический уровень повышается в ответ на генотоксический или экзогенный стресс. Потенциально, дефицит в аутофагии может способствовать более высокой уязвимости и более длительный период восстановления нейронов обычно наблюдается при травматической травме головного мозга и повреждении спинного мозга. Кроме того, раковые стволовые клетки ЦНС, такие как стволовые клетки глиобластомы, часто характеризуются изменениями в аутофагическом потоке.
Связанная с ЦНС аутофагия
Аутофагия - процесс катаболической переработки клеточных компонентов - была в центре внимания при лечении опухолей, связанных с ЦНС, и поврежденных нервов.
Митофагия

Митохондрии, двухмембранные органеллы, выполняют основные метаболические функции, включающие биосинтез АТФ через окислительное фосфорилирование, поддержание производства активных видов кислорода, активацию апоптоза и гомеостаз кальция. Как только повреждение митохондрий становится слишком сильным, дисфункциональные митохондрии не могут слиться с нормальными соседними митохондриями. Аутофагический поток как процесс самоедания включает в себя образование аутофагосом, слияние аутофагосом с лизосомами и деградацию аутолизосом. Потенциально нарушенный аутофагический поток может повлиять на ликвидацию поврежденных митохондрий. Аналогично, по сравнению со здоровыми митохондриями, поврежденные митохондрии могут производить больше ROS для повышения выживаемости опухолевых клеток. Митофагия оказывает проинвазивное воздействие на клетки глиобластомы. Галавотти создали новый механизм, с помощью которого связанный с аутофагией фактор P62 и DRAM-1 контролирует миграцию и инвазию в стволовые клетки глиобластомы (GSCs) посредством регуляции коэффициента потребления кислорода (OCR) - индекса активности транспортной электронной цепи в митохондриях.
Ферритинофагия/ферроптоз
Ядерный рецептор-коактиватор 4 и эндогенная световая цепь 3B связывают ферритин в аутофагосомах, что вызывает ферритинофагию для разрушения внутриклеточного ферритина. Кроме того, ферритинофагия может вызвать ферроптоз. Недавний эксперимент подтвердил гипотезу о том, что ферроптоз способствует развитию PD и дофаминергических нейронов за счет активации белковой киназы Cα. Кроме того, домен HECT и RCC1-подобный домен 2 действуют как ключевой регулятор ферритинофагии и способствуют возникновению и прогрессированию ряда нейрогенетических заболеваний. Что касается опухолей ЦНС, активация ферритинофагии может также тормозить прогрессирование и злокачественность опухолей, таких как глиомы.