Название этой работы показалось мне очень многообещающим и я решил в нее заглубиться. К сожалению, не все нашел, что хотелось бы. Но, для любителей спортивной физиологии, возможно, найдутся интересные моменты.
Снова на повестке феномен так называемого "повторного спринта". От каких физиологических механизмов зависит способность многократно повторять усилия высокой интенсивности?
Ответ на этот вопрос пробуют найти авторы сегодняшней работы. Изучение этой проблемы представляется важным для таких видов спорта, как спортивные игры и единоборства.

Недавно мы говорили о том, как влияет дополнительный кислород на повторные спринты:
Так же у нас была работа о влиянии ограничения кровотока на восстановление между спринтами:
Сегодня же спринты с измерениями лактата, закисления, креатинфосфата и других параметров.
Идея исследования
Оценить связь работоспособности при выполнении повторного спринта с изменениями концентрации АТФ, креатинфосфата и ионов водорода в мышцах, а также с нейро-мышечной активностью.
Испытуемые
8 мужчин спортсменов-любителей командных видов спорта (19,5±0,9 лет, 183,4±5,4 см, 80,3±8,7 кг и МПК 53,9±5,8 мл/кг/мин) приняли участие в этом исследовании.
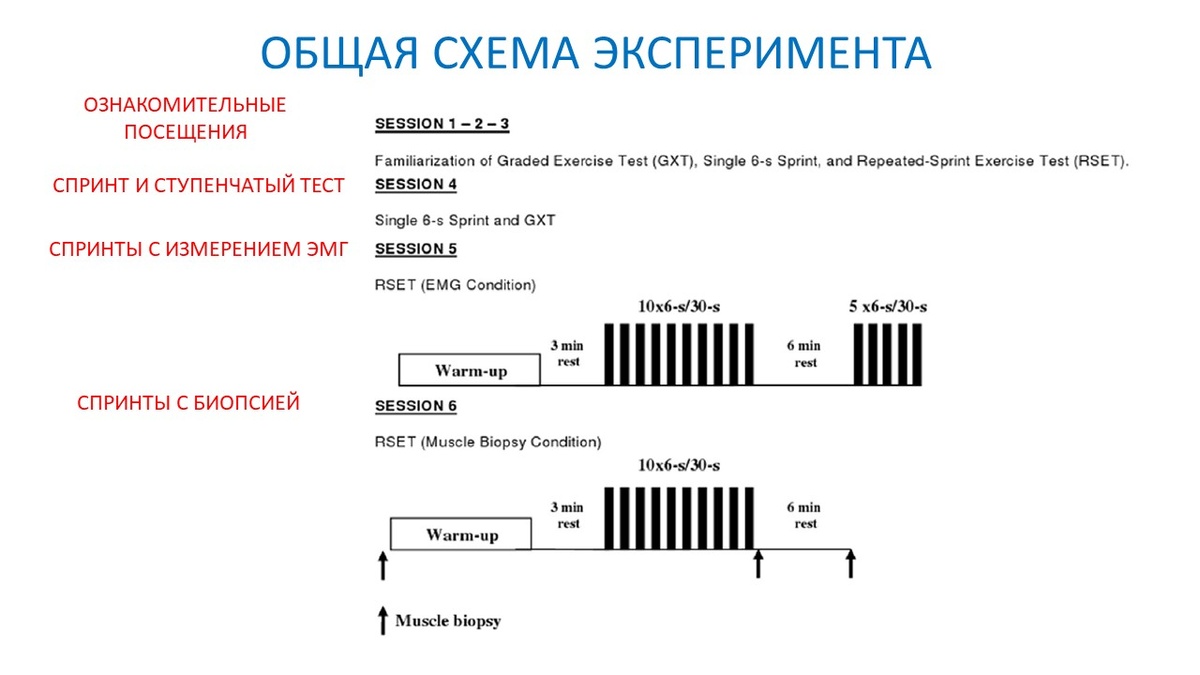
Дизайн эксперимента
- Испытуемые посетили лабораторию 6 раз: 1-3 предварительные процедуры, 4-6 экспериментальные дни (далее их описание);
- День 4: испытуемые выполнили один 6-секундный спринт "во всю" на велоэргометре. После 30 минут отдыха выполнялся тест с возрастающей нагрузкой для определения показателей выносливости (рис. 1);
- День 5: испытуемые выполнили десять 6-секундных спринтов на велоэргометре с отдыхом 30 секунд между ними. Далее, после 6 минут пассивного отдыха, выполнили еще пять таких же спринтов (рис. 1);
- День 6: испытуемые также выполнили десять 6-секундных спринтов, но работа уже сопровождалась взятием проб мышечной ткани (биопсии) в покое, сразу после 10 спринтов и через 6 минут восстановления (рис. 1).
Что измеряли
- Максимальные характеристики одного спринта;
- В первый день повторных спринтов - электрическую активность мышцы бедра;
- Во второй день повторных спринтов - изменение мышечных метаболитов.
Результаты
Изменения параметров работоспособности
- Значения пиковой мощности (ПМ) и общей выполненной работы (ОВР) показаны на рисунке 2;
- Уменьшение ПМ и ОВР после первых 5 спринтов в среднем составило 13,8±5,7% и 17,1±6,1% соответственно;
- Самые высокие значения ПМ и ОВР наблюдались во время первого спринта и были значительно выше, чем в любом из других 14 спринтов;
- За первые 10 спринтов средние значения ПМ и ОВР снизились на 24,1±9,4% и 27,2±9,4% от максимального значения (спринт 1) соответственно;
- Во время 11 спринта (после 6 минут отдыха) значения ПМ и ОВР значительно восстановились по сравнению с показателями 10 спринта, но остались на 12,8±6,1% и 13,7±7,7% ниже значений максимального спринта;
- За последние 5 спринтов (с 11 по 15) ПМ и ОВР снова снизились на 17±8,7% и 20,3±9,4% соответственно (рис. 2);
- Во время последнего спринта ПМ и ОВР были на 27,3±11,5% и 30,8±12,4% ниже значений максимального спринта;
- Соотношение общей работы за 5 последовательных спринтов к работе одного первого спринта в спринтах 11 и 15 было значительно ниже значений, полученных для спринтов 1 и 5 (Рис. 3);
- Это соотношение использовалось в качестве индекса работоспособности при повторных спринтах;
Электрическая активность мышц
- Значения электрической активности мышц (амплитуды ЭМГ) при повторных спринтах показаны на рисунке 4;
- По сравнению с 1-м спринтом снижение мышечной активности после первых пяти спринтов в среднем составило 9,2±6,2%;
- За первые 10 спринтов значение мышечной активности снизилось уже на 14,3±7,6%;
- После 6 минут отдыха во время 11 спринта мышечная активность не показала никакого восстановления после спринта 10 и была на 18,3±12,4% ниже начальных значений (рис. 4);
- Во время последующих спринтов (с 11 по 15) наблюдалось дальнейшее значительное снижение ЭМГ-сигнала;
- Значения ЭМГ для спринта 15 были на 4,4±5,4% и 22,1±11% ниже значений, достигнутых в спринте 11 и спринте 1 соответственно.
Изменения мышечных метаболитов и показателя pH
- На рисунке 5 показаны средние концентрации некоторых мышечных метаболитов и значения pH до, сразу после 10 спринтов и после 6 минут восстановления;
- После спринта 10 наблюдалось снижение содержания АТФ на 29,5±9,6%;
- После 6-минутного восстановления АТФ была ресинтезирована до 92,6±6,% от значения покоя;
- Содержание креатинфосфата в мышцах после спринта 10 составило 46,9±21,4% от значения покоя;
- Креатинфосфат был восстановлен до 85,3±10,3% к 6-й минуте отдыха, что оставалось все еще значительно ниже значения покоя;
- Содержание лактата в мышцах увеличилось на 921±232% после 10-го спринта, а затем значительно снизилось до 559±184% после 6-минутного отдыха;
- Концентрация ионов водорода в мышцах увеличилась до 191±50% после 10-го спринта, а после 6-минутного отдыха снизилась до 144±32% от значений покоя;
- Показатель pH в мышце оставался сниженным по сравнению со значениями покоя после 6-минутного отдыха (рис. 5);
- На рисунке 6 показано отношение общей выполненной работы (ОВР) к мышечной активности (МА, т.е. нейромышечная эффективность) во время двух серий спринтов;
- Отношение ОВР/МА, выраженное в процентах от значения первого спринта, снизилось в 5-м спринте (- 8,6±7,1%) и в 10-м спринте (- 15,1±13,6%) по отношению к исходному значению в спринте 1;
- 6-минутный отдых увеличил отношение ОВР/МА в 11-м спринте, которое было значительно больше, чем в спринте 10 (+20.7±8.5%) и незначительно больше, чем в спринте 1 (рис. 6);
- Значения для спринта 15 были на 16,8±7,6% ниже значений, достигнутых в спринте 11.
Связь между переменными
- Ресинтез креатинфосфата после 6 мин отдыха положительно коррелировал с общей работой в спринте 11 (r = 0,79) и работой в пяти спринтах (т.е. в спринтах 11-15, r = 0,67, рис. 7);
- Никаких корреляций не было обнаружено между какими-либо переменными работоспособности во второй серии спринтов и изменениями в каком-либо из мышечных метаболитов;
- Также не было обнаружено значимых корреляций между восстановлением работоспособности и изменением электрической активности мышцы.
Выводы
- Наблюдалось значительное снижение концентрации как АТФ и креатинфосфата в мышцах, так и активности двигательных единиц мышц во время выполнения повторных спринтов;
- Восстановление работоспособности в повторных спринтах коррелировало с ресинтезом креатинфосфата, но не с удалением ионов водорода;
- Активность двигательных единиц оставалась сниженной даже после отдыха после первых 10 спринтов;
- Работоспособность в повторных спринтах после отдыха восстанавливалась в меньшей степени, чем мощность и работа одиночного спринта;
- Более низкая мощность работы во время спринтов после 6-минутного отдыха, скорее всего, была связана с внутримышечными, а не с нейронными факторами.
P.S. К сожалению, авторами не приводится взаимосвязь показателей выносливости (МПК, м.б. ПАНО) со способностью выполнять повторные спринты. Хотя тест этот они проводили(!) в день 4 (рис. 1). Вот это было бы интересно. Может быть оставили материал для другой статьи, такое бывает.
Также для понимания физиологии и биоэнергетики упражнения очень полезны цифры с динамикой энергетических фосфатов и других метаболитов. Теперь осталось понять какой механизм будет ответственен за снижение скорости расхода креатинфосфата и за повышение скорости его восстановления. Аэробные возможности мышц? Может быть. Продолжим потихоньку изучать тему повторных спринтов.
Получить доступ к неопубликованным лекциям, статьям и другим материалам, а также поддержать нашу работу можно на Boosty
Сейчас там опубликованы следующие материалы:
1. Конкурентный тренинг (лекция).
2. Весогонка в единоборствах (лекция).
3. Плиометрика в фитнесе (лекция).
4. Метаболические эффекты упражнений (доклад).
5. Зона жиросжигания - что мы о ней знаем (лекция).
6. Физиология аэробного и анаэробного порогов, МПК (лекция).
7. Все про Дроп-Сет (лекция).
8. Локальное жиросжигание (лекция).
9. Ишемический тренинг. Часть 1 и 2 (научный доклад).
10. Физподготовка в волейболе (три лекции).
11. Биоэнергетика спорта (обзорная лекция).
12. Классификация аэробных и анаэробных упражнений (лекция).
13. Статодинамика: 30 лет обсуждений Часть 1 (лекция).
14. Семинар «Физиология жиросжигания» (3 лекции по 3 часа).
15. Разное количество повторений. Обзор эффектов для массы, силы и выносливости (лекция).
16. Предпосылки для локального жиросжигания (доклад).
17. Концепция физподготовки на примере игрового вида спорта (лекция).
18. Теория и методика интервальной тренировки (лекция 2 часа).
19. Некоторые экспериментальные факты о физической подготовке боксера (доклад).
20. Дефицит калорий: диета или упражнение? (лекция).
21. NEW! Влияние физкультуры на здоровье. Новые научные данные (лекция).
22. NEW! Теория и методика силовой тренировки. (Лекция. Часть 1 и 2).
Дополнительные материалы по этой теме:
Вклад различных энергетических систем при выполнении коротких максимальных упражнений
Работа сердца при выполнении повторных спринтерских ускорений
Энергообеспечение высокоинтенсивной интервальной тренировки
Метаболизм спринтерской интервальной тренировки
Источник: Mendez-Villanueva A, Edge J, Suriano R, Hamer P, Bishop D. The recovery of repeated-sprint exercise is associated with PCr resynthesis, while muscle pH and EMG amplitude remain depressed. PLoS One. 2012;7(12):e51977.