При чтении материалов данного раздела учебника по транскрипции можно наблюдать следующие положения.
Первое. Если при описании репликации расплетению молекулы уделяется большое внимание – говориться и о топоизомеразах и хеликазе, то при описании режима транскрипции о таких элементах в явном виде ничего не говорится и поэтому даже не понятно расплетается ли транскрибируемый участок молекулы или нет. Во многих других материалах описывается процесс расплетения транскрибируемого гена при помощи топоизомераз, а вот в учебнике это все как-то завуалировано. Хотя учебник довольно нов, он переиздан в 2012 году.
Второе. При транскрипции, как и во всех остальных случаях, в предыдущих материалах, ничего не говорится о хиральности, как будь то она и вовсе лишняя и ни на что не влияет.
Почему так? На молекулярном уровне знаний это приемлемо, так как в этом случае эти положения транскрипции можно объяснить либо ложно, либо вообще ничего не объяснять. На квантовом уровне познания эти явления возможно описать более верно. Естественно молекулярный уровень указывает на многие истинные положения транскрипции и многие другие положения молекулярной биологии. Вот пример:
“Гены разных гистонов сцеплены. Все они транскрибируются по отдельности, часто с противоположных цепей ДНК. В геноме гистоновые гены распределены неодинаково. У одних видов они образуют тандемные повторы, у птиц и млекопитающих (рис. 87) рассеяны по всему геному. Большинство гистоновых генов лишено интронов, а у мРНК отсутствуют 3'-поли(А)-концы”. (Стр. 167).
На удивление в данной цитате верны все утверждения, кроме утверждения транскрибируются. Это потому, что ученым не понятно, как же вообще происходит транскрибирование и что именно запускает режим транскрипции. Вот режим репликации совершенно очевидно запускается тепловыми фотонами. Это абсолютно необходимое условие, хотя иногда и недостаточное, например, семена без влаги не произрастают. А что запускает транскрипцию? Считается, что сразу надо найти какую-то последовательность предшествующую транскрибируемого гена и затем уже начать работать с ним. Как отыскивать требуемый ген наука не знает. Но допустим ген найден, тогда современная молекулярная биология предполагает этот ген надо расплести и затем на расплетенной цепи построить РНК, которую после этого отсоединить от матрицы, на которой дальше будет построена молекула белка.
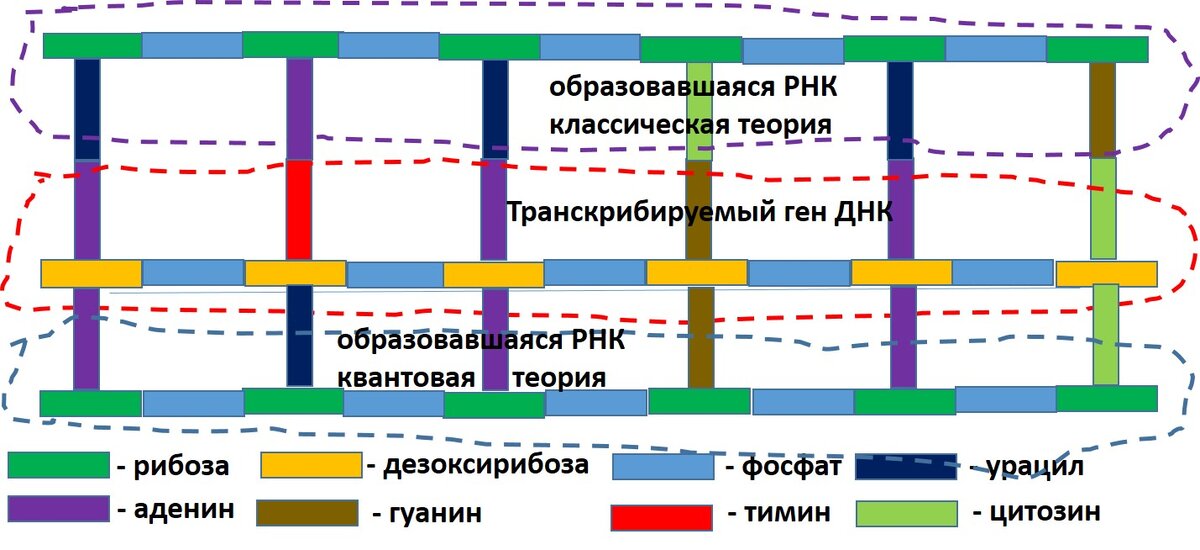
На этом рисунке изображен транскрибируемый ген. Согласно классической теории на нем настраивается комплементарная цепь, как и при репликации, только вместо тимина работает урацил и вместо дезоксирибозы используется сахар рибоза. Уже этим все кажется странным. Обычно (при репликации) на расплетенном участке ДНК сразу же начинает строиться дочерняя цепь ДНК, а тут вдруг оказывается, что на аденине, которому при репликации требуется тинин или нуклеотид с тинином, должен строиться урацил с рибозой. Кто или что должен решить, что в данном случае настраивать на аденине?
Но самим главным непонятным и никак не объясняемым явлением в транскрипции – это экспрессия генов. Предположим, что у каждого гена есть свой личный признак, а без этого
природе, никак не обойтись. Таких признаков должно быть около 25 000, по числу работающих генов. Каждый ген должен транскрибироваться в определенном месте и в определенное время. Перестановка генов местами или изменение времени их экспрессии приводит к гибели организма.
Все описываемые в учебнике “–азы” и другие подобные элементы находятся внутри клетки, как они определяют, когда и какой ген надо экспрессировать? Сами они не могут это знать, даже если они могут что-то узнавать, куда-то двигаться, что-то вырезать, сшивать и так далее. Для этого им должно быть дано указание из вне клетки. И таким сигналом может быть только морфологическое указание: вот здесь запустите транскрипцию актина, а вот здесь запустите меелин и так далее.
А морфологическим сигналом может быть только электрический ток (электрический ток – это поток фотонов). Например, у человека растет в длину и толщину нога. Массив мяса (актина и миозина) нарастает в процессе синтеза этих белков в клетках. Как клетка, точнее ее органеллы, знают, что надо в это время в этом месте строить актин или миозин? Синтез этих или других белков в этом случае не может происходить без четкой организации, иначе получится какая-нибудь слоновость (ноги разбухают, как у слона) или другая неприятность.
Есть несколько моделей морфогенеза, и клеточный, и химический, и еще какие-то, но проникновение в клетку морфогенических атомов, молекул или даже каких-то частиц весьма сомнительно. Да и самое главное откуда им взяться? А фотону это сделать просто. Как формируются эти фотоны рассказано в статье “Морфогенез”.
Факт это, то, что та или иная клетка, а, следовательно, и все ее органеллы попадают под это специфическое излучение. И это специфическое излучение должно запустить режим транскрипции именно нужного в этой точке белка. Как это можно сделать?
Первое это нужно активизировать, например, какую-нибудь полимеразу. А что значит активизировать? Это значить заставить ее что-то совершать с молекулой, но для этого этой полимеразе надо передать информацию о гене, с которым следует работать. Главное передать название этого гена. Если допустим идентификатор гена закодирован в его ТАТА-боксе или в какой-то другой последовательности, то следует этот код передать поисковику полимеразы или другой органеллы, начинающей работать с именно требуемым геном или кластером, согласно учебнику. Это есть начало экспрессии гена.
Поисковик как-то находит указанный ген и включает в работу следующие элементы полимеразы и чего-то другого. Дальше, согласно классической теории, требуемый ген расплетается и на одной из цепей ДНК строится цепь РНК. Верхняя часть рисунка.
Квантовая теория предполагает, что ничего в природе этого нет. В самом деле все происходит так.
Действительно существует морфологическое поле – это обычное всем известное голографическое поле, создаваемое фотонами различной энергии. Эти фотоны и экспрессируют тот или иной ген, в том или другом месте. Но воздействуют они не на полимеразы или подобные им элементы, а непосредственно на электроны самого гена. Для этого не нужны никакие кэпы, ни полимеразы, ни белковые факторы транскрипции, ни ТАТА-боксы, ни что другое, так красочно расписанное в учебнике и вбиваемое в головы студентов.
У человека примерно 25 000 генов, которые следует запускать на транскрипцию, следовательно, столько же требуется и запускающих сигналов как минимум. Большое число сигналов генерируется при других процессах в организме: при репликации, при трансляции, митозе и так далее. И эти все сигналы вырабатываются параллельно сигналам морфогенеза. Чтобы эти сигналы не накладывались друг на друга природа разделила их по качеству. Сигналы морфогенеза она сделала правыми (см. статьи о кванте). Все фотоны, создающие морфологическое строение организма в виде голографии, обладают правой ориентации, в отличие от всех остальных фотонов организма.
По этой причине в ДНК присутствует правая дезоксирибоза. Ее некоторые электроны способны излучать и поглощать фотоны правой ориентации, а большинство других электронов этой же молекулы поглощают и излучают фотоны левой ориентации. Оказывается, что морфологическое (голографическое) поле создается фотонами правой ориентации (хиральности). Если ДНК попадает под какое-то излучение этого поля, то, соответственно, величине энергии этого поля возбуждается (экспрессируется) требуемый ген. Для возбуждения каждого гена требуется своя энергия. Почему так получилось в природе, я сказать не могу, точнее, у меня нет убедительной модели для самого себя. Предполагаю, что это эволюционное явление.
Это значит, что хиральный узел начинает светится таким светом, который становится привлекательным для аденина, или урацила, или гуанина, или цитозина. Причем хиральный узел, к которому уже присоединен аденин (в составе молекулы) требует присоединения (такова энергия фотонов) к этому узлу аденина или нуклеотида с аденином. Но вполне может быть, что и в этом случае соблюдается комплементарность. Узел с тимином требует связи с урацилом, узел с гуанином требует гуанин, тоже и с цитозином.
Так строиться копия гена без расплетения молекулы.
Наука знает, что некоторые аминокислоты также обладают хиральностью. При загрузке аминокислоты азотистыми основаниями, получается транспортная РНК. На ней находится триплет, соответствующий этой аминокислоте. Если свет триплета возбужденного гена будет привлекателен для триплета тРНК, соответствующей аминокислоты, то к ДНК присоединяться не нуклеотиды мРНК, а триплеты тРНК. И таким образом на гене ДНК синтезируется на РНК, а сразу синтезируется белок.
Именно так синтезируются все гистоны, тубулин и все другие внутриклеточные белки. Каждая клетка знает, когда и какой ей требуется белок. А вот морфологические белки синтезируются через РНК.
Как мы знаем белок способен сворачиваться в глобулы. В третичной структуре молекула белка образует множество водородных связей между различными радикалами аминокислот, составляющих молекулу. Это, примерно, как клейкая лента, которая все время пытается слипнуться какими-нибудь местами. Иногда белки сворачиваются в регулярные формы. Например, тубулин сворачивается в виде элементов микротрубочек.
Если в глобулу, или любую другую пространственную структуру, сворачивается несколько типов белка, то такая структура называется четвертичной.
Вот такие структуры и образуют гистоны. Не молекула обворачивается вокруг группы белков, а белки гистоны, сворачиваясь, тянут за собой молекулу. В результате окажется, что вокруг гистонов будет обвита только та часть молекулы, на которой уже не возможна транскрипция. А те части, на которых можно производить транскрипцию, остаются свободными. И в этом случае молекулу при транскрипции не нужно скручивать с гистонов и расплетать.
Но гистоны служат не только для конденсации молекулы, а еще и служат для сортировки репликационных молекул при митозе. Клеточные центры снабжены микротрубочками, которые образуют так называемое веретено деления. Похоже, что у каждого центра свои микротрубочки. Одни микротрубочки состоят из α-тубулина, а другие микротрубочки состоят из β-тубулина. По этой причине один клеточный центр тянет к себе гравитационно одну цепь молекулы за гистоновые белки, а другой центр тянет к себе вторую цепь молекулы. Гистоновые белки на разных цепях оказываются не симметричными и одни белки оказываются резонансными для α-тубулина, а другие для β-тубулина.
Немножко об этих проблемах можно прочесть в статьях “Полчаса митоза” и “Тройка, шестерка, код”.



















