Сложно, сложно, не понятно…
Эта статья во многом основана на замечательной книге Евгения Кунина «Логика случая», в которой вы сможете найти более подробное обсуждение затронутых здесь вопросов. В частности, в этой статье были опущены почти все математические выкладки.
Идея этой статьи родилась из следующего видимого парадокса. Как все прекрасно знают, когда-то живые организмы все были одноклеточными, потом появились многоклеточные губки, медузы и прочие примитивные животные, из которых ещё позднее развились позвоночные, которые вышли на сушу, превратились в амфибий, рептилий, млекопитающих и наконец в человека. Всё выглядит так, будто эволюционная история — это движение от простого к сложному, от примитивного к продвинутому.

Однако, наверное, любой, кто начинает заниматься биологией, вскоре обнаруживает, что очень часто дело обстоит иначе: сложное превращается в простое. Посмотрим на это дело с позиции зоологии беспозвоночных. При изучении любой группы животных начинают обычно со свободноживущих сложных форм, а затем переходят ко всякого рода паразитам и симбионтам, которые обычно устроены значительно проще. При этом часто очень хорошо прослеживается утрата различных органов или даже целых систем органов при переходе к другому образу жизни. В первую очередь обычно утрачиваются органы чувств, так как видеть или слышать внутри другого животного особо нечего, затем значительно редуцируется нервная система, что связано как с упрощением поведения, так и с утратой большей части сигналов от внешнего мира. Далее может значительно упрощаться или вовсе утрачиваться пищеварительная система и т.д. Например, в школьной биологии это всё легко наблюдать на плоских червях.
Приведём другой пример, не связанный с зоологией — посмотрим на происхождение эукариот. Ещё в конце прошлого века было установлено, что ближайшими родственниками эукариот являются археи, а не бактерии. Сравнение геномов показывает, что большая часть генов, отвечающих за репликацию, репарацию, транскрипцию, трансляцию, протеолиз, деградацию РНК и значительная часть генов, отвечающих за строение хроматина, РНК-интерференцию и прочее имеют архейное происхождение. Однако любопытный факт заключается в том, что все эти гены встречаются в разных комбинациях у разных архей, и можно заключить возможность двух сценариев: или эукариоты получили все эти гены в результате длительного процесса горизонтального переноса или, что более эволюционно экономно, большая часть этих генов уже присутствовала у архейного предка эукариот. Во втором случае мы можем допустить существование значительно большей сложности древних архей по сравнению с современными. Однако это ещё не всё. Одним из главных отличительных признаков эукариот является наличие митохондрий — внутриклеточных симбионтов, потомков бактерий. Однако у некоторых одноклеточных эукариот митохондрий нет, например, у представителей группы Parabasalia.
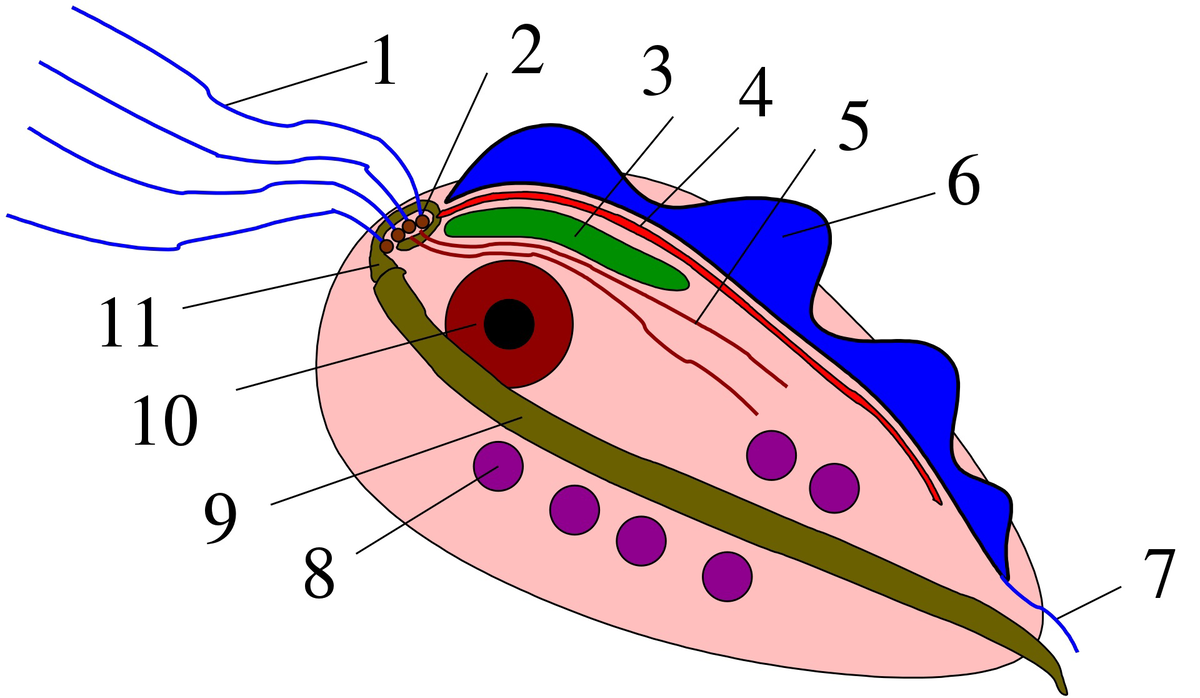
Логично было предположить, что и было сделано учёными, что хотя бы часть безмитохондриальных эукариот никогда их не имела, а эндосимбиоз с бактериями произошёл позже отделения этих групп. Однако позднее во всех без исключения безмитохондриальных эукариотах были найдены различные тельца, являющиеся сильно редуцированными за ненадобностью митохондриями. Например, у организма на картинке это гидрогеносомы под цифрой 8. Из всего вышесказанного следует, что предок всех современных эукариот был не только потомком каких-то древних сложных архей, имевших огромный набор генов, но и уже имел митохондрии, которые дальше могли только упрощаться.
Обратимся наконец к происхождению жизни. Все самые простые свободноживущие организмы являются очень сложными: они имеют клеточное строение, тысячи генов, все основные метаболические пути и т.д. Все самые простые организмы: вирусы и паразитические бактерии опять же являются паразитами и как минимум во многих случаях — результатом упрощения. Никаких переходных форм между не клетками и клетками мы не наблюдаем.
Приведённые выше примеры иллюстрируют общую тенденцию: мы легко можем проследить историю упрощения, но почти никогда не видим, каким образом организмы в процессе эволюции усложняются. Мы видим червей на разных стадиях утраты органов, но не видим или почти не видим их на разных стадиях приобретения новых органов. Мы видим эукариот с редуцированными митохондриями, но не видим эукариот, никогда митохондрий не имевших. Видим современных простых архей, но не видим их вероятно более сложного предка или по крайней мере не можем проследить историю усложнения до уровня предка эукариот и т.д. Это всё подводит нас к неожиданному выводу: эволюция в целом идёт по пути упрощения.
Если эволюция идёт по пути упрощения, то откуда взялись такие сложные мы? Чтобы ответить на этот вопрос, для начала договоримся о терминах. Термин сложность кажется не очень строгим, однако его можно формализовать в терминах теории информации и термодинамики. Сложным является объект, для описания которого нужно много информации, или иными словами — система с высокой энтропией. Согласно определению Шеннона, энтропия сообщения (описания системы) вычисляется по формуле:
где p — вероятности каждого из n разных исходов, или в случае сообщения — вероятность встречи конкретного символа на данной позиции. Поясним на примере сразу из биологии. Пусть наше сообщение — это последовательность ДНК. Последовательность только аденинов будет иметь энтропию равную 0, так как вероятность встретить A на каждом сайте равна 1, а log1=0.
AAAAAAAAAAAAAAAAAAAAAAAAA
Если в последовательности с равной вероятностью встречаются все нуклеотида, то её энтропия будет максимальной
ACACGTATTGCCAAGATGACGAATGCCA
Мы видим, что такое определение согласуется с термодинамическим пониманием энтропии: для наиболее хаотичной последовательности энтропия имеет наибольшее значение. Существенно, что энтропия сообщения зависит от того, какие вероятности для каждой буквы мы определим. То есть вопрос состоит в том, чтобы таким образом определить вероятность появления для каждого нуклеотида, чтобы посчитанное нами значение имело наибольший биологический смысл. Для этого было предложено использовать частоту встречи нуклеотида на данной позиции в выравнивании гомологичных последовательностей. Иными словами, если какой-то участок генома очень консервативен (похож у разных организмов), его энтропия будет маленькой, а если он сильно варьируется, то энтропия будет большой. Далее для каждой позиции в геноме посчитаем её информационное содержание. Математические детали мы опустим, отметив только, что если энтропия позиции больше, информации в ней содержится меньше. Посчитав информационное содержание для каждого сайта в геноме и разделив на общее число нуклеотидов, мы получим плотность информации. Чем больше плотность информации, тем меньше энтропия генома.
Понятно, что в реальной жизни все подобные расчёты будут очень приблизительными, однако грубую оценку дать они способны. Результаты расчётов для нескольких организмов приведены на рисунке
Мы видим, что у организмов, которые мы обычно считаем более сложными (человек, дрозофила), плотность информации значительно ниже, чем у простых вирусов и бактерий. То есть энтропия геномов сложных организмов выше. С одной стороны это не удивительно для любого человека, знакомого с тем фактом, что у нас большая часть генома не кодирует вообще ничего, в то время как у бактерий гены расположены очень плотно, и геном в целом утроен гораздо экономнее. С другой стороны мы смогли формализовать связь между сложностью и плотностью информации, и теперь можем сделать такой вывод: геномы сложных организмов всегда устроены неэкономно, то есть в них много «мусора». Как выяснится позднее, этот вывод будет ключом к разрешению «парадокса сложности».
Чтобы найти следующую зацепку для наших рассуждений, обратим внимание на ещё один интересный факт: в эволюционной истории нашего вида почти всегда были только самые «ущербные» организмы. Этот тезис во многом является оценочным, поэтому доказать строго его нельзя, но можно проиллюстрировать примерами. Археи вытеснены бактериями почти из всех мест обитания, одножгутиковые протисты сильно уступают по разноорбразию прочим, вторичноротые распространены гораздо меньше первичноротых, приматы — далеко не самая процветающая группа млекопитающих и т.д. Мы как будто всегда были на обочине эволюционной истории, и у этого есть своё объяснение.
Чтобы понять последующие рассуждения, немного поговорим о популяционной генетике. В этой области биологии существует такое правило: только мутации, для которых |s|»1/N могут быть эффективно зафиксированы или отбракованы естественным отбором. Здесь s — коэффициент отбора (разница в приспособленности организма с мутацией и без), N — эффективный размер популяции. Из этой формулы следует, что в очень маленькой популяции будут фиксироваться только очень полезные мутации, а отбраковываться только очень вредные.
Допустим, у нас есть большая популяция, и в ней произошла какая-то относительно вредная мутация. Поскольку эта мутация снижает приспособленность, она вскоре исчезнет из популяции в следствие естественного отбора. Однако если популяция маленькая, эта мутация может очень долго отбором не вымываться, так как в маленьких популяциях по понятным причинам большую роль играет случай — так называемый дрейф генов. Итак, у организмов с небольшой численностью особей, в геноме будет накапливаться много умеренно вредных мутаций, которые потом однако могут стать материалом для последующего отбора.
Мы говорили ранее, что у сложных организмов в геномах много разных некодирующих последовательностей. Такой геномный «мусор» вообще говоря снижает приспособленность, так как требует энергетических затрат на репликацию, уменьшает скорость репликации, усложняет регуляцию и т.д. Однако с другой стороны именно эта ненужная поначалу ДНК может стать источником для новых генов, то есть для последующего усложнения организма. Таким образом низкая плотность информации генома является важнейшей предпосылкой для формирования сложности. А накопиться такие изменения из-за своей вредности могут только в популяциях с небольшим эффективным размером. Это по видимому обязательное условие, но конечно же не достаточное, ведь скорее всего такая популяция попросту вымрет под грузом накопившихся геномных изменений. Усложнение — сценарий маловероятный, но возможный. Получается, что мы такие сложные из-за того, что среди наших предков никогда не было процветающих организмов. Наших предков постоянно теснили, их популяции были маленькими, что создавало необходимые условия для усложнения организации.
Вот мы и подошли к ответу на парадокс биологической сложности. Эволюция действительно преимущественно работает на упрощение. Это легко объяснимо: упрощаться проще, чем усложняться. Однако в небольших популяциях создаются условия для накопления умеренно вредных мутаций, которые затем могут послужить материалом для скачка на другой уровень организации. Такие скачки не являются преобладающим эволюционным процессом, а являются в каком-то смысле статистическими выбросами. Если мы мысленно построим распределение организмов по сложности, то большая часть из них расположится в районе простых и несложных форм, однако у этого распределения будет также и длинный хвост справа: где-то на конце него и расположимся мы.