
Происхождение разнообразия и устойчивости СК
Введение
Накопленный материал по динамике поведения стволовых клеток (СК), с одной стороны: предполагает сложные механизмы управления их свойствами и их трансформациями, с другой: требует простых общих принципов организации поведения СК. Иначе, невозможно представить настолько надёжно и точно работающие механизмы онтогенеза, восстановления и самовозобновления организма. Представляется необходимым ввести ряд постулатов, для которых есть основания в виде наблюдаемых процессов и накопленных представлений о таковых. Они касаются общей модели поведения СК в организме. И, как представляется автору, помогут закрепить рамки поисков целей, в исследованиях СК, в продуктивных направлениях, чтобы оптимизировать их и существенно сократить расходы на исследования. С другой стороны: определить исключения и также определить модель их реализации.
Основная часть
Большое разнообразие СК в организме морфологически - эволюционно продвинутых организмов (теплокровных, млекопитающих..) ставит серьёзные задачи по обобщению новых генетических данных и замыкания их в теоретические контуры, которые бы включали в себя и генезис их происхождения. Во-первых: чтобы найти недостающие звенья цепи генетических преобразований при развитии организма.
Во-вторых: чтобы найти генетические принципы организации и динамического развития органов и организма.
В-третьих: чтобы выяснить существование скрытой патологии в развитии органов и соединительных тканей.
В-четвёртых: чтобы найти цепочки маркерных признаков, позволяющих достоверно определять типы СК, а значит и функционирование тканей в динамике их развития.
Ясно, что существует жёсткая логическая схема динамики развития организма и его органов. Вряд ли природа создала много вариантов развития таких «проектов». Мы начнём с главных действующих лиц в этой саге о строительстве творения более грандиозного - чем пирамида Хеопса и даже наша Солнечная Система. Наглядные примеры, которые бы подтверждали наши модели, мы будем брать и из области патологий, таких как онкология, так как их процессы – суть «снимки» с элементов реальных процессов, только выполненные не ко времени и с ошибками. Эволюция иерархических отношений СК совпадает с эволюцией усложнения организмов (7,8) и было бы нелогичным ожидать иного источника происхождения иерархической модели СК. Усложнение иерархии СК связано и сопровождается усложнением организмов, как и функционирования органообразования в организмах. Вопросы эволюции гетерогенности СК ждут своего Колумба. В то же время стоит сразу отделить два аспекта эволюции : организма(как сложноклеточного сообщества) и генома организма (как генотипа).
СК – пока они представляют из себя «черные ящики» в изобилии разбросанных по организму. Но, всё же, составляющих ничтожную часть клеточного гетеронаселения организма. Примерно на 10^11 клеток в организме человека присутствуют около 10^9 СК, причем, сначала, основная масса их находится в костном мозге, с возрастом наблюдается серьёзное истощение этого ресурса. Другой ресурс – тканевые островки, эндотелий, лимфатические узлы, тестис, тонкие слои в почках, главном мозге. Основное деление СК на Эмбриональные (ЭК), Фетальные(ФК) и Постнатальные(ПК). ЭК и ФК – считаются плюропотентными, то есть, способными стать родоначальниками любых других видов клеток. ПК – уже представляют из себя сложноорганизованную группу СК. В данной работе мы будем исследовать всю иерархию СК, поэтому, изначально, мы стираем этот переход и предполагаем непрерывность трансформации и генерации видов СК, что естественно и логично. Таким образом, с более высоким уровнем СК – плюропотентность их снижается, вырождаясь у прогениторов до выбора 2-3 типов дифференцированных клеток (ДК).
См. Рис 1 Иерархия СК.
Это первый постулат – все СК организма образуют единую иерархию непрерывных взаимных трансформаций, в нашей гипотезе о динамике трансформации СК в организме. Получается, что динамика развития будет непрерывной и все изменения в организме будут последовательными "архитектурными" перестройками и надстройками, а не «революционными» процессами. Таким образом Рис 1, образуется иерархия СК(1,2,). Здесь появится необходимость ещё в нескольких постулатах. Так, нужно ответить на вопрос: как организм сохраняет свою организационную стабильность, почему так надёжно работают механизмы восстановления эпителия, соединительной ткани, костей и др.? Необходимо предположить, что СК (прогениторы) - которые обслуживают данный участок ткани – получают точный сигнал о необходимой замене. Т.е., через плотный контакт СК с ДК и через экзосомы (4-5) Рис 2.
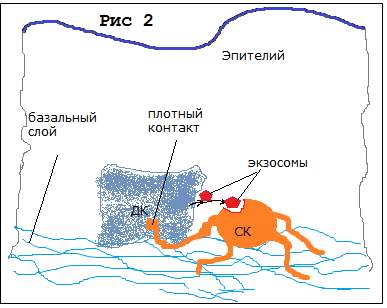
Рис 2 Схема обмена информацией через экзосомы или контакт между ДК и СК
Это будет второй постулат – Наличие обмена информацией между СК и клетками ДК через экзосомы и плотные контакты, который достаточно просто объяснит, почему так точно восстанавливаются быстро повреждаемые ткани организма. СК – получают сигнал от клетки, которая исчерпала временной или физический ресурс (1,3,4,6,7). В этом сигнале содержится информация о типе клетки и прогенитор узнает, во что он должен трансформироваться (как ассиметричным делением на себя самого и нужный тип клетки, так и просто трансформацией (например прогениторов нейронов и астроцитов).
Третий постулат- трансформации СК в органах и лимфоузлах: для нормального выполнения второго постулата, СК в реальном организме,
Км – стволовые клетки костного мозга.
ОТ – Области Трансформации .
Рис 3. Схема движения СК через центры трансформации в различных органах.
«испытывают последовательные трансформации» в лимфоузлах и органах организма. Т.о., на место своей «работы» СК подходят подготовленными – Рис 3. Поэтому, тут вызывают вопросы предлагаемые взгляды на плюоропотентность и индуцирование её. Неустойчивость их поведения (дифференциация в разные типы клеток) – признак неестественности течения событий(5).
Четвёртый постулат – наличие Ниши для СК низкого типа, который следует из третьего: Для работы СК - нужна «НИША», т.е. место, где биохимические условия достаточно стабильны и присутствуют «соседи», необходимые для выполнения операций активирования деления и правильной дифференциации в нужную СК (4-6). Более того, для нормального функционирования такой ниши там должна быть ещё одна СК. Именно симбиоз двух типов СК устойчиво даст третий тип СК (делением более высокого типа СК), результатом будет появление СК нового типа. Например (Рис 4): (СК5 + СК6) - образуют «нишу», после деления получаем – (СК5 + СК6 + СК7) где: СК7 - новый тип, эта СК покидает нишу (Рис4).
Рис 4 СК 5(А5) и СК 6(А6) в нише делится СК 6 новая СК 7 (А7) покидает нишу.
Так работают СК в костном мозге, в желудочках Головного Мозга (ГМ) и в половых тканях организмов. Прогениторы могут делиться и дифференцироваться «по ходу». Тогда работает механизм плотного контакта, поглощения прогенитором экзосомы или через «временной» таймер запуска, когда состав окружающей среды и время – являются факторами влияющими на дифференциацию прогенитора. Последний механизм не является точным, поэтому могут наступать локальные дисгармонии в архитектуре органа (эпителия) – он определяет возрастные изменения кожи и органов (и гетеронаселение при раке). Тут нужно заметить, что в начале эмбрионального развития организуется уникальная ситуация – ниши совмещены топологически - ТСН. Поэтому, в эмбрионе организуются два разных типа регионов, где ещё продолжается ТСН и где включается уже обычный механизм – топологического разделения ниш - ТРН.
Обсуждение
Написанные постулаты прямо следуют из накопленного фактологического материала. Они не противоречиво объединяют неудачи манипуляций с СК разного типа, так и манипуляции с индуцированными iСК (иСК). Тут нужно обратиться к еще одной стороне вопроса: топологией клеток. Автором предполагается следующая примерная топология клетки рис 5,рис 6. Предлагается вертикальная организация доменов клетки. Т.е., все продукты управления (ПУ): белки, микроРНК и месенджРНК и двигаются по оси ядро-поры ядра- цитоплазма-мембрана клетки. Причем, основными путями служит актин – образуя линейные структуры, которые имеют строго направленную организацию потоков ПУ. Такая структура очень хорошо решает проблему высокой специфичности и эффективности управления и ответов в клетках. Очень похоже на то, что в доменах есть восходящая часть потоков и нисходящая часть потоков рис7 , в этих потоках движется цитоплазма с восстановленным и окисленными продуктами, органеллы и продукты (гормоны, продукты синтеза клетки и т.п.. , это бы в тысячи раз снизило энергозатраты на транспорт ПУ в клетке. О потоках внутри клетки упоминается в большом количестве исследований, в обзоре 10 приводятся данные об установленных цитоплазматических потоках как между клетками, так и внутри клеток. Более того, видимо, существуют потоки с высокими [O] [pH] и низкими [O] [pH] –рис 7. А это огромная доля от всех энергозатрат в клетке (где-то более 90%). Учитывая это, заметим, что реорганизация клетки – будет серьёзно детерминирована предшествующим состоянием клетки.
Трудно представить себе, чтобы клетки имели полные карты трансформаций из одного произвольного состояния в другое произвольное состояние (типы клеток) «архитектурного» состояния – в любое другое. Тут мы находим поддержку Первому Постулату в этой статье. На рис 5 дано «лицо» типа клетки, которое может быть легко определено различными маркерами (флуоресцентными). При этом, можно сохранять клетки неповрежденными но – эффективно и с высокой специфичностью отсортированными и зафиксированными.
Рис 5 Картография клетки (в координатах 00-00, 00-15,180-180 и т.д. - это рецепторы, каналы и сигнальные точки клетки)
Рис 6 Центростремительная организация клетки.
На Рис 6 представлена схема организации домена клетки от гетерохроматина ядра до соответствующего ему участка внешней мембраны клетки.
Рис 7 Направление микропотоков внутри клетки.
Заключение
На рисунках представлены модели организации топологии клеток. Как видно, они сегодня не противоречат нашим представлениям о работе клетки. В то же время они позволяют понять причину эффективности работы транспортной системы клетки, а также дают новое представлгние о дополнительных явлениях сопровождающих процессинг клетки.
Библиография
1.Self-formationoffunctionaladenohypophysisinthree-dimensionalcultureHidetakaSuga, TaisukeKadoshima, MakiMinaguchi, MasatoshiOhgushi, MikaSoen, TokushigeNakano, NozomuTakata, TakafumiWataya, KeikoMuguruma, HiroyukiMiyoshi, ShigenobuYonemura, YutakaOiso & YoshikiSasaiNature (2011)
Publishedonline 09 November 2011 –( Уэтой работы ,кроме прекрасной приложимости ктончайшему восстановлению врегенеративной терапии мозга ииспользования всего современного инструментария висследованиях, есть ещё одна сторона. Это ещё одно наглядное подтверждение модели иерархии стволовых клеток в маленьком регионе при организации гипофиза. Эта модель давно была дана в нескольких ресурсах (2006г -7г). Основой её служит непосредственный контакт двух типов СК. Который порождает третий вид. Тоже было обнаружено недавно и в «нишах» костного мозга, Тоже было открыто и в иерархии Самых первых эмбриональных клеток . Т.о., Модель полностью подтверждена ,сверху донизу в иерархиях ответвлений разных популяций СК.).
2.Unravelling cancer stem cell potential
..Benjamin Beck & Cédric Blanpain
Nature Reviews Cancer 13, 727–738 (2013) doi:10.1038/nrc3597
Published online 24 September 2013". (подтверждаетсянаонкомоделях)
3.Wnt activity and basal niche position sensitize intestinal stem and progenitor cells to DNA damage
Si Tao, Duozhuang Tang, Yohei Morita, Tobias Sperka, Omid Omrani, André Lechel,Vadim Sakk, Johann Kraus, Hans A Kestler, Michael Kühl, Karl Lenhard Rudolph
The EMBO Journal (2015) 34, 624-640
4.A family business: stem cell progeny join the niche to regulate homeostasis
Ya-Chieh Hsu1 & Elaine Fuchs1, Nature Reviews Molecular Cell Biology 13, 103-114 (February 2012)
5.A niche role for cancer exosomes in metastasisYun Zhang & Xiao-Fan Wang
Nature Cell Biology 17,709–711 (2015) doi:10.1038/ncb3181 Published online 29 May 2015
6.Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver
Bruno Costa-Silva, Nicole M. Aiello, Allyson J. Ocean,
et al Nature Cell Biology 17, 816–826 (2015)
7.The establishment of Caenorhabditis elegans germline pattern is controlled by overlapping proximal and distal somatic gonad signals.
Pepper AS, Lo TW, Killian DJ, Hall DH, Hubbard EJ.
Dev Biol. 2003;259:336–350. [PubMed]
8.Male and female Drosophila germline stem cells: two versions of immortality.
Fuller MT, Spradling AC Science.2007;316:402–404.
9.Massive expression of germ cell-specific genes is a hallmark of cancer and a potential target for novel treatment development , Bruggeman JW 1 , Костер J 2 , Лоддер P 1 , Реппинг S 1 , Хамер G 3 , Oncogene. 2018 Oct;37(42):5694-5700. doi: 10.1038/s41388-018-0357-2.
10. Intercellular cytoplasm transport during Drosophila oogenesis. Mahajan-Miklos S1, Cooley L. Dev. Biol. 165,336–351 1994
--
Последние работы по топологии цитологии:
Long-range interactions between topologically associating domains shape the four-dimensional genome during differentiationJonas Paulsen, Tharvesh M. Liyakat Ali, Maxim Nekrasov, Erwan Delbarre, Marie-Odile Baudement, Sebastian Kurscheid, David Tremethick & Philippe CollasNature Geneticsvolume 51, pages835–843 (2019)



















