Традиционно считается, что эволюция направляется "слепым" естественным отбором, где после случайных мутаций выживают наиболее приспособленные. В заметке описана модель, из которой следует, что главную роль играет "зрячий" половой отбор, который пытается заполнить пространство нейтральных к выживанию признаков. Из разумного предположения, что новые экологические ниши находятся вдалеке от имеющихся, следует, что в большинстве случаев эволюция направляется половым отбором, а не естественным. Что хорошо согласуется с эмпирическими данными, в которых без полового отбора эволюция ничего сложнее микроба не создала.
Происходит это по той же причине, по которой подброшенная 100 раз монетка не может выпасть орлом существенно далеко от 50. В то время как половой отбор, используя всего одну эвристику - "ищи похожего на себя", собирает доступные гены, действующие в направлении от центра распределения. Модель отличается от большинства общеизвестных генетических моделей, тем что рассматривается сумма генов, а не один-два важных гена, функция полового отбора - наиболее схожая по внешнему признаку особь, и функции выживания в самой модели нет.
Составим модель из:
а) 1000 особей, каждая из которых имеет 100 генов с нулевым или единичным аллелем
б) у каждой особи есть внешне наблюдаемый признак, которые равен сумме значений его генов, то есть принимает значения от 0 до 100
в) нулевое поколение создано из особей, где вероятность нулевого и единичного аллеля в каждом гене одинакова
Рассмотрим два варианта порождения следующего поколения:
г) бесполый, где каждая особь дает одного потомка в котором случайно выбираемый один из генов меняется на противоположный
д) двуполой, где наиболее близкие по значению признака особи собираются в пары, и каждая пара порождает два потомка, у которых каждый ген случайно выбран из соответствующих родительских генов, а также у каждого потомка один из генов меняется на противоположный
Поскольку вероятность мутации каждого гена одинакова, в более частые аллели мутации легче попасть. Для особи с 60 единичными генами вероятность мутации единичного гена в нулевой равна 0.6, а обратная равна 0.4. Что создает "тягу к центру" для потомков выдающихся особей. И, наоборот, при половом размножении за одно поколение потомки выдающихся особей 100 раз (по числу выбираемых у родителей генов) с выгодной вероятностью пытаются отойти от центра и только в одном гене случайно мутируют с невыгодной вероятностью. Другими словами, в бесполой популяции в конкурсе на самого выдающегося потомка участвуют только один-два самых выдающихся родителей с края, а в двуполой популяции участвует вся элита. Для дальнейшего расхождения двуполой популяции от центра важно, что неудачные потомки выдающихся родителей замещаются удачными потомками менее выдающихся родителей.
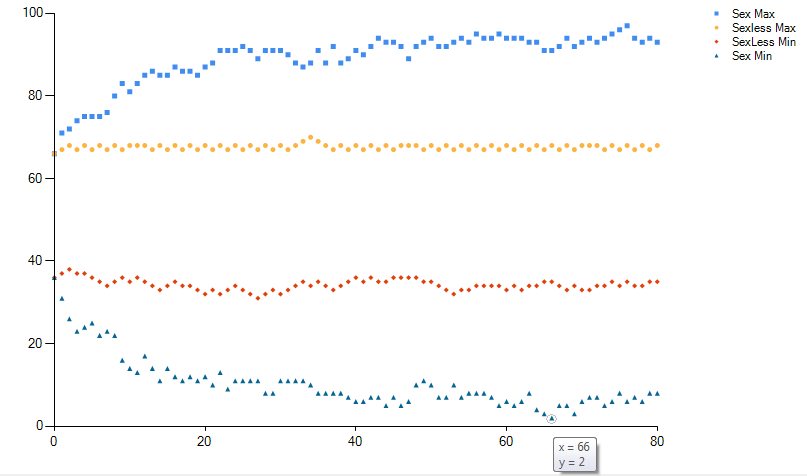
Один из вариантов компьютерной симуляции показан на рисунке. Самые выдающиеся особи из бесполой популяции доходят до 30 и до 70, но не могут пройти дальше к 20 и 80. А в двуполой популяции одна особь дошла до минимального значения 2 за 66 поколений, а максимальное значение 97 было достигнуто за 76 поколений. С математической точки зрения бесполая популяция оверфитится, без необходимости слишком сильно "прижимаясь" к центру распределения. А половое размножение выступает в роли регуляризатора, "разжимая" популяцию по доступному нейтральному пространству. Тем самым приспосабливаясь не только к прошлым и текущим оптимумам, а к будущим и неизвестным.
Бесполая популяция из-за "вероятностного барьера" :
а) не переживет существенное изменение нейтральности признака, если для выживания потребуются особи далеко от центра распределения (в нашем примере это область ниже 20 и выше 80)
б) не может выйти на следующую экологическую нишу если эта ниша расположена далеко от центра распределения (строго говоря если экологические ниши расположены в одном и том же центре логичней их считать одной нишей)
в) не может дособирать редкие появляющиеся гены для увеличения нейтрального пространства
г) не может ожидать на краю распределения если проход в экологическую нишу открывается время от времени
д) не может пройти цепочку экологических ниш (сложная эволюция), если хотя бы один переход расположен далеко от центра распределения.
Из пункта д) следует запрет на сложную эволюцию для бесполого размножения. Это запрет статистического характера. Всегда есть вероятность, что молекулы сами соберутся в автомобиль. Но она настолько низкая, что при виде автомобиля мы понимаем, что его кто-то создал. Сложная эволюция, в том виде, как мы её знаем, создана половым отбором.
[1] https://en.wikipedia.org/wiki/Sexual_selection