
Появление в Европе предков современных индоевропейцев - носителей Y-ДНК или отцовских гаплогрупп R1a и R1b произошло несколько тысячелетий назад и, вероятно, в разное время и по совершенно разным направлениям. Почему есть такое предположение? Во-первых, это связано с самыми древними носителями данных гаплогрупп, проникших в определенное время в Европу, а, во-вторых, с аутосомными примесями, которые различаются у индивидов-носителей R1a и R1b. Какая есть на этот счет информация, полученная из научных исследований?
Начнем с R1a. Известно, что наиболее древние носители гаплогруппы R1a в Европе обнаружены в северных регионах нынешней России, а самая древняя известная R1a (субклад R1a1b~-YP1272>YP1301) определена у представителя культуры Веретье (10785—10626 лет до н. э.) из могильника Песчаница у озера Лача в Каргопольском районе Архангельской области (Lehti Saag et al. 2021). Другая древняя R1a1 определена у охотника с Южного Оленьего острова на Онежском озере, жившего примерно 8 тыс. лет назад (W.Haak et al., 2015). Все это указывает на то, что где-то в промежутке между 13-14 тысячами лет назад охотники гаплогруппы R1a направились на север Сибири вслед за животными, на которых они охотились. Это было связано с резким потеплением Арктики и отступлением ледников. Спустя какое-то время сибирские охотники-собиратели достигли северных районов Европы. Примерно в это же время другая группа сибирских охотников (носители гаплогруппы Q1) также устремилась на север, но повернули в другую сторону - на восток, и примерно в те же сроки (13-14 тысяч лет назад) достигли побережья Северной Америки.
Условия в арктической мамонтовой степи тогда были существенно другими - на Крайнем Севере было значительно теплее, чем сейчас. Так, на востоке Сибири белоствольные берёзы встречались в то время вплоть до широты современного Северного Ледовитого океана. Сейчас эти деревья растут значительно южнее, а современные условия намного суровее, чем те, что были там 13 тысячелетий назад. Новое похолодание в Арктике наступило значительно позже, после чего лес вновь сместился к югу, а в Арктике опять образовалась тундра, которую мы и наблюдаем сейчас. Вполне возможно, что процесс миграции носителей R1a в Европу продолжался много тысячелетий, в том числе и более южными путями после последовавшего на Крайнем Севере очередного сильного похолодания. На это косвенно могут указывать некоторые факты: 1) на севере России найдены пока представители только архаичных субкладов R1a, которые сейчас в Европе практически не встречаются 2) в Сибири древняя R1a1-M17 была определена у индивида из Иркутской области (Глазковский некрополь) возрастом 8000-7000 лет назад, что указывает на то, что еще в эпоху Неолита в Сибири оставались архаичные носители этой гаплогруппы, которые могли продолжать миграции в Восточную Европу, но более южными путями. По аутосомным показателям R1a относились к восточно-европейским охотниками-собирателями (EHG). Главный предковый компонент их связан с племенами древних северных евразийцев (ANE), заселивших в Мезолите северную и восточную Европу. В лесных и лесо-степных районах Русской равнины в эпоху Неолита часть восточных охотников-собирателей EHG смешивались с племенами западных охотников-собирателей (WHG), которые были вытеснены ранними и более поздними неолитическими земледельцами на восток и север Европы. Во времена Неолита и Бронзового века на севере и в центре Русской равнины преобладали три отцовские линии - R1a (большинство), I2a и Q1, давшие позднее начало культуре Боевых топоров (или Шнуровой керамики), а в южно-русских степях преобладалт R1b и в явно меньшей степени I2a, которые потом дали начало степной Ямной культуре.
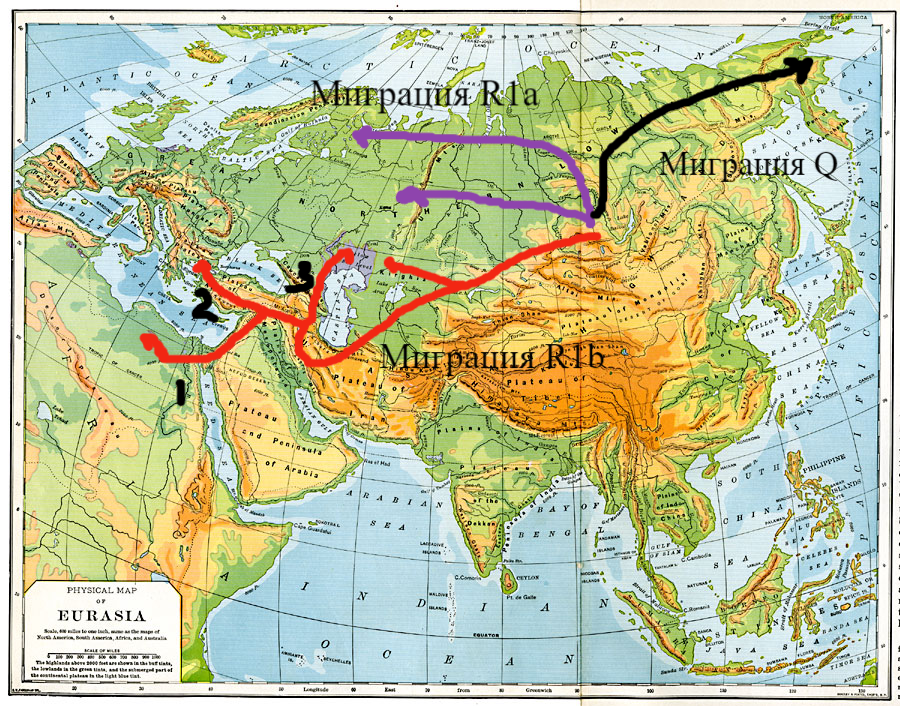
Теперь остановимся на племенах-носителях Y-хромосомеой гаплогруппы R1b. Известно, что R1b отделились от общего ствола предков R1 где-то в Южной Сибири или Средней Азии. Далее они направились на юг и достигли районов "плодородного полумесяца" на Ближнем Востоке. Все три их основные ветви R1b1 (R1b1a, R1b1b, R1b1c), вероятно, произошли именно там. Самая южная из ветвей R1b1c (V88), встречается в основном в Леванте и Африке (на рисунке №1 она обозначена цифрой 1). Вторая ветвь R1b1b (M335) была обнаружена только в Анатолии (на рисунке выше обозначена цифрой 2). Третья или северная ветвь, R1b1a (P297), вероятно, возникла южнее Кавказа - в Иране, восточной Анатолии или северной Месопотамии, а затем уже пересекла Кавказ и проникла в южно-русские степи (на рисунке №1 она обозначена цифрой 3), откуда позже одна ее часть вторглась в Центральную и Западную Европу, а другая - в Среднюю Азию и Сибирь. Предполагается, что третья ветвь R1b1a (P297), вероятно, пересекла Кавказ вдоль западного побережья Каспийского моря по пути ассимилировав местное население. Это подтверждают недавние полногеномные исследования, которые показали, что Ямная культура, располагавшаяся в южно-русских степях, несмотря на значительное преобладание в ней отцовской линии R1b и в явно меньшей степени I2a, является результатом смешения разных популяций людей: мужчин из восточных и западных охотников-собирателей (EHG и WHG), с одной стороны, и женщин из кавказских охотников-собирателей (CHG), анатолийских, иранских и левантийских неолитических фермеров (EEF), с другой стороны. Это смешение популяций произошло где-то около 4000 г. до н.э. (Lazaridis et al., 2022; Allentoft et al., 2022; Chintalapati et al., 2022). Процент EHG к остальным предковым компонетам Ямной культуры был в районе 50%, что указывает на то, что в этой миграции принимали участие в основном мужчины, а женщин они брали из числа покоренных народов Кавказа, Ирана, Анатолии или Ближнего Востока. Таким образом, проникновение племен R1b в Восточную Европу было значительно позднее (на 5-7 тысячелетий), чем племен-носителей R1a.
Возможно, что некоторая часть носителей R1b мигрировали на Южный Урал и Понтийско-Каспийскую степь напрямую из Южной Сибири или Средней Азии. На это, в частности, указывает генетический состав Хвалынской культуры на средней и южной Волге. Эта культура была предшедственницей Ямной культуры и в ней также явно преобладала отцовская Y-ДНК R1b, но почти отсутствовали предковые компоненты (определены по аутосомным показателям) из Анатолии, Леванта и Ирана-Кавказа, которые найдены в Ямной культуре.
Почему многие западные ученые выводят культуру Шнуровой Керамики (КШК) из Ямной культуры, хотя преобладающие мужские линии Y-ДНК у них сильно различались (R1a и R1b соответственно)? Все дело в том, что обе эти культуры несли высокий процент (не менее 50%) предковых компонентов, полученных от древних северных евразийцев (ANE), а также проникшие в обе культуры геномы мигрантов с Запада - племен западных охотников-собирателей (WHG) с основной отцовской линией I2a. Парадоксальность этого несоответствия и его ошибочность впервые отметил авторитетный российский археолог С.Клейн, который вступил в дискуссию с известными западными палеогенетиками. Он, в частности, писал: "Во-первых, с точки зрения археологии эти две культуры не являются родственными, так как захоронения у шнуровиков (КШК) очень часто бескурганные, а погребенные лежат в иных позах (на боку), чем у ямников (на спине с поднятыми коленами), мужчины иначе, чем женщины, а их керамика совершенно иная, вооружение у КШК – каменные боевые топоры и молоты, а у ямников - молоточковидные булавки, а также медные копьевидные ножи или шилья. Кроме того, все погребения у ямников обильно посыпаны охрой, а у КШК этого нет. Во-вторых, язык, на котором говорило население Ямной ку-ры, не мог быть протоиндоевропейским, т.к. разница во времени между распадом протоиндоевропейского языка и распадом ямной культуры слишком велика.
В-третьих, крайне странно распределен степной генетический вклад среди шнуровиков и родственных им культур (Haak et al. 2015). Очень богатый на Севере Европы и все менее и менее выраженный по мере приближения к югу, где как раз находится основной ареал культуры ямников. Такое распределение степного компонента представляется непонятным, если считать, что источник вклада в геном шнуровиков – это ямная культура на юго-востоке России, но это же распределение будет вполне естественным, если предположить, что общий источник геномного сходства находится не возле ямной культуры, а на Севере Европы. Данные показывает, что "ямный" генетический компонент – вряд ли на самом деле "ямный" по своему происхождению. Скорее всего этот компонент появился у населения Северной Европы, а уже оттуда он распространился как в степь, так и в другие культуры Европы".
С Клейном нельзя не согласится. Западные палеогенетики не учли как раз тот самый предковый компонент ANE древних северных евразийцев, больше всего распространенный среди EHG (восточно-европейских охотников-собирателей). Этот предковый компонент в Европе довольно быстро убывает с севера на юг. По этой причине, происхождение КШК (культуры шнуровой керамики или же боевых топоров с доминированием R1a) и Ямной культуры с доминированием R1b), скорее всего, независимы друг от друга.
Если Вам понравилась статья подписывайтесь на канал, делитесь ею в социальных сетях и ставьте лайк (👍 - палец вверх)
Читайте также другие наши статьи: