Монофилия - это одно из центральных понятий в филогенетике, и оно принципиально важно для интерпретации полученных результатов. Естественная группа с общей эволюционной историей - это по сути монофилетическая группа. И это то, что мы пытаемся понять, является ли изучаемая группа естественной или нет.
Я здесь следую трактовке Хеннига, и даю определения согласно кладистической парадигме.
Монофилетической называется группа, которая включает предка и всех его потомков. Монофилетическая группа это синоним термина "клада", потому что клада по определению монофилетична.
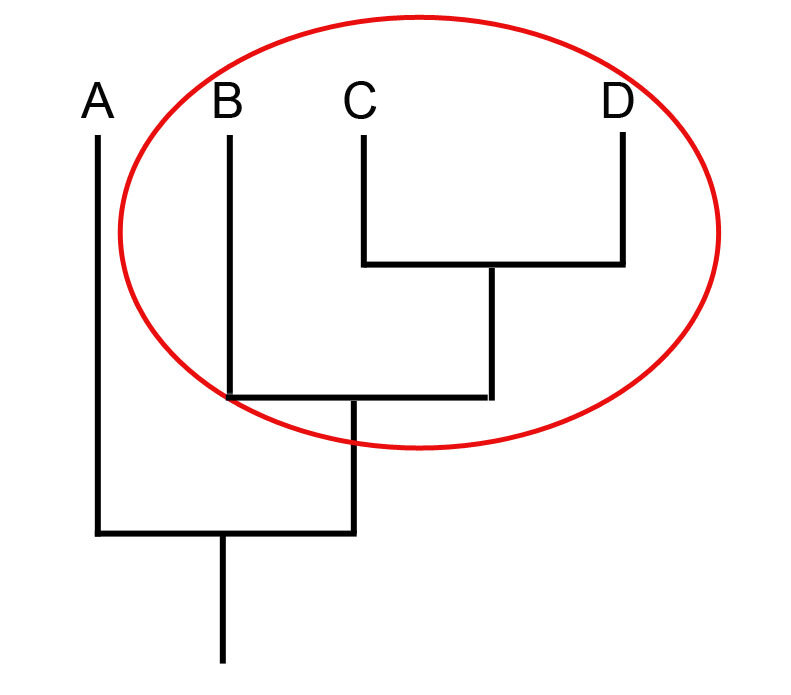
Сестринская группа - это группа, с которой изучаемый таксон формирует кладу (или монофилию). Например, таксон D сестринский по отношению к таксону С, справедливо и обратное. А вот таксон B сестринский по отношению к кладе, которая сформирована C и D, и он равноудален от этих двух таксонов.
На филогении имеет смысл обсуждать только монофилетические группы. Если группа немонофилетическая, как правило, о ней мало что можно сказать, кроме установления этого факта.
Еще есть путаница с интерпретацией филогений. Если два таксона расположены физически рядом на дереве, интуитивно хочется сделать вывод, что они родственны. Однако если они не формируют кладу, то это не так, Рассмотрим пример на картинке ниже. Я специально выбрала путающую форму записи, но тем не менее она также правильная:
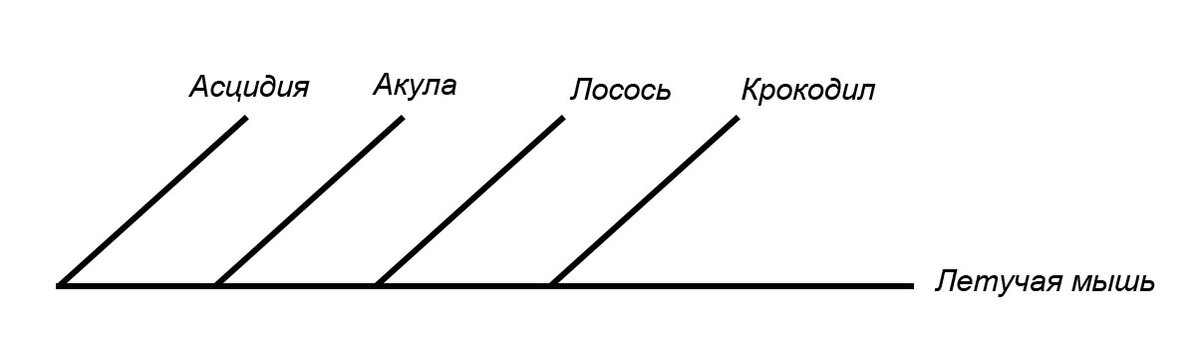
На этой записи лосось и акула рядом, и хочется сделать вывод, что они ближе, чем, к примеру, акула и летучая мышь. Но это было бы неправильно. Здесь лосось, крокодил и летучая мышь формируют кладу, а акула - сестринская группа для этой клады. Акула одинакова далека от всех таксонов внутри этой клады.
Парафилетическая группа включает предка и не всех его потомков. Например, долгое время термиты считались отдельным отрядом. Уже давно было показано, что это такие специализированные тараканы, и, соответственно, отряд Таракановые стал парафилетическим, потому что термитов он не включал. Сейчас их объединяют в один отряд (Inward et al. 2007, Legendre et al. 2015).
Самый известный пример - это рептилии и птицы. Раньше считалось, что птицы и рептилии - это отдельные группы, но потом выяснилось, что птицы - это такая специализированная ветвь динозавров, то есть они произошли от рептилий и по сути ими являются. Таким образом, рептилии без птиц - это парафилетическая группа (Agnolin et al. 2019).
Еще один подобные пример - это копытные. Еще в конце 20го века было показано, что китообразные произошли от них, и, соответственно, копытные без китообразных - это парафилетический таксон. Сейчас их всех объединяют в отряд Китопарнокопытных (Zurano et al. 2019).
Полифилетическая группа не включает общего предка. То есть по
сути какая-то "сборная солянка", объединение таксонов, не родственных друг другу. Например, раньше панголинов и трубкозубов выделяли в отдельный отряд Edentata вместе. Потом выяснилось, что панголины ближе к Хищным, а трубкозубы входят в кладу афротерий, которая среди прочих включает слонов и морских коров.
Есть небольшая путаница в названиях. В классическом понимании эволюционных биологов монофилетический таксон разделяется на голофилетический и парафилетический. Голофилетический при этом соответствует монофилетическому по Хеннигу. Логика тут такая, монофилетические таксоны в классическом понимании это любые естественные таксоны, и парафилетические таксоны тоже естественные, потому что все таксоны происходят от других таксонов. Например, представим себе, что у нас есть вид на материке, и одна его популяция попала каким-то образом на остров. На острове она быстро эволюционировала из-за дрейфа генов и стала существенно отличаться от своего материкового предка, но материковый вид при этом мало изменился. Получается, что материковый вид - это парафилетический таксон, потому что не включает своего потомка (островной вид).
Это вполне резонно. Но все же, во-первых, такое встречается не очень часто. Обычно все же мы имеем дело с уже существенно разошедшимися генетически видами. Во-вторых, это обычно имеет смыл только на уровне видов, если действительно есть две сущности, которые не обмениваются генетическим материалом. И то, если они генетическим материалом не обмениваются уже какое-то время, то скорее всего на кладограмме они все же будут формировать разные клады. Но если мы покажем, например, что какой-то род произошел от другого рода, то нам лучше синонимизировать их. Иначе нам не избежать вопроса: "Насколько должна быть существенной разница, чтобы выделять парафилетическую группу? Где провести эту грань?" Какой-то вид в роде сильнее отличается от других. Можем ли мы на этом основании описать новый род? В таксономии такой подход приведет к неразберихе.
Вторая проблема с классической интерпретацией естественных групп в том, что на практике оказывается, что монофилетические группы по Хеннигу обычно интерпретируются довольно хорошо по крайней мере в случае хороших поддержек, а вот парафилетические и полифилетические группы иногда сложно отличить друг от друга, просто потому что не всегда можно сказать, каким был предок. В случае монофилетического таксона, если он включает все таксоны в кладе, не очень важно, каким был предок в основании этой клады. Для того, чтобы назвать таксон пара- или полифилетическим это бывает принципиально важно.
Приведу в пример свою работу. Филогения и ревизия трибы клопов-слепняков Monaloniini (Namyatova & Cassis 2016).
Я привожу свою филогению из статьи. Суть в том, что изначально были две трибы - Monaloniini (на филогении отмечены голубым) и Odoniellini (на филогении отмечены коричневым). У меня получилось, что роды не формируют этих триб, и вместе перемешиваются, так что пришлось свести в синонимы Odoniellini к Monaloniini. На картинке клада, которая объединяет все роды (Monaloniini в новом понимании), обозначена красным кружком.
Но вопрос в том, эти трибы в прошлом их понимании пара- или полифилетические? В основании филогении клада, которая сформирована родами, которые ранее были как в Monaloniini (Arculanus и Mansoniella), так и в Odoniellini (Chamus). Для того, чтобы считать Monaloniini в прежнем понимании монофилетической группой, нам надо показать, что общий предок был Monaloniini, но мы этого не знаем. Тоже самое справедливо для Odoniellini. А учитывая, что поддержки в основании небольшие, то обсуждение этих вопросов вообще не очень осмысленно. Мы четко видим на кладограмме, что эти две трибы немонофилетичны, а какие они (пара- или полифилические) - не понятно. Но если подумать, так ли это важно, если все равно мы будем теперь называть все эти роды Monaloniini? В статье я аккуратно писала про трибы в прежнем понимании как non-monophyletic.
Кстати, именно поддержки часто являются фактором, который не дает с большой вероятностью отнести таксон к поли- или парафилетическому. Да что уж там, даже монофилия иногда по этой причине под вопросом. Поэтому действительно, иногда лучше говорить о том, что таксон вероятно монофилетический или немонофилетический (в понимании Хеннига), а изучение того, как точно расположены таксоны на эволюционном древе в определенной группе иногда занимает много лет, и даже геномика не всегда дает однозначный ответ.
Еще раз хочу обратить ваше внимание на относительность, о которой уже говорила применительно к признакам. Будет ли у нас подтверждена монофилия, которая реально существует, зависит от качества данных и все. Потому что если таксон - это клада, хорошие данные в итоге это покажут. К примеру, из приведенной выше филогении можно исключить любой таксон или группу таксонов, Monaloniini в новом понимании останутся монофилетическими.
Однако будет ли таксон пара- и полифилетическим также зависит от набора таксонов в нашем исследовании. В своей работе по Monaloniini мне удалось включить все роды в филогению, что не всегда бывает возможным. Допустим, род Chamus не был бы описан, или был бы таким редким, что я бы не включила его в исследование. В таком случае, можно было бы сделать вывод что общей предок всей группы принадлежит Monaloniini, таким образом Odoniellini в прежнем понимании был бы монофилетической группой внутри парафилетической трибы Monaloniini.
Agnolin, F. L., Motta, M. J., Brissón Egli, F., Lo Coco, G., & Novas, F. E. (2019). Paravian phylogeny and the dinosaur-bird transition: an overview. Frontiers in Earth Science, 6, 252.
Inward, D., Beccaloni, G., & Eggleton, P. (2007). Death of an order: a comprehensive molecular phylogenetic study confirms that termites are eusocial cockroaches. Biology letters, 3(3), 331-335.
Legendre, F., Nel, A., Svenson, G. J., Robillard, T., Pellens, R., & Grandcolas, P. (2015). Phylogeny of Dictyoptera: dating the origin of cockroaches, praying mantises and termites with molecular data and controlled fossil evidence. Plos one, 10(7), e0130127.
Namyatova, A. A., & Cassis, G. (2016). Systematic revision and phylogeny of the plant bug tribe Monaloniini (Insecta: Heteroptera: Miridae: Bryocorinae) of the world. Zoological Journal of the Linnean Society, 176(1), 36-136.
Zurano, J. P., Magalhães, F. M., Asato, A. E., Silva, G., Bidau, C. J., Mesquita, D. O., & Costa, G. C. (2019). Cetartiodactyla: updating a time-calibrated molecular phylogeny. Molecular phylogenetics and evolution, 133, 256-262.