Основная фабрика по производству коллагена – это фибробласты. Напомню, что молекула коллагена состоит из 3 альфа-цепей, каждая из которых может кодироваться собственным геном, поэтому существует так много разновидностей коллагена. Лучше всего изучены синтез и сборка коллагена I типа, в организме человека эта разновидность коллагена встречается чаще других типов.
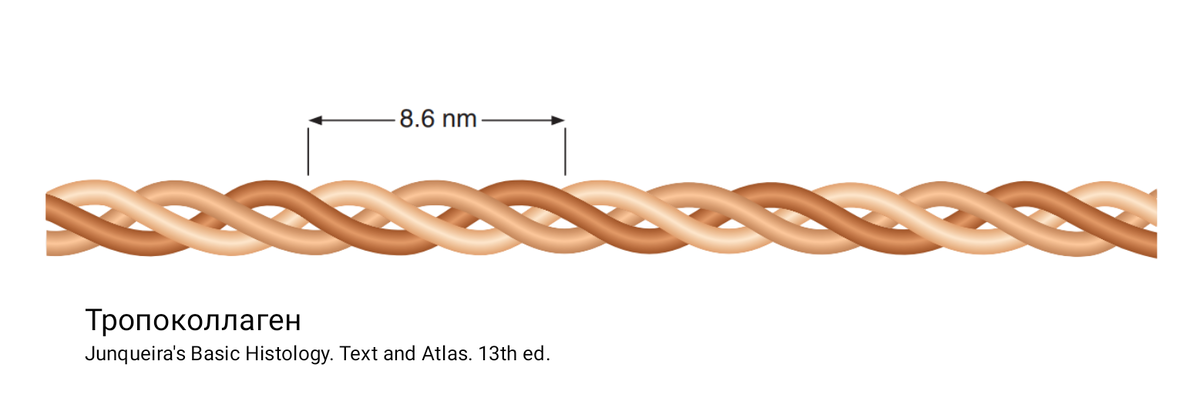
Любознательные учёные даже попытались определить скорость синтеза коллагена. Они вводили животным меченный тритием пролин (пролин – это самая характерная аминокислота коллагена).
Уже через 3 минуты 83% меченного пролина связывалось с эндоплазматической сетью, а 5% пролина доходило до комплекса Гольджи. Через 20 минут почти весь пролин скапливался в области комплекса Гольджи, а затем появлялся на поверхности фибробласта. Как видите, клетка делает всё быстро, ведь коллаген – это материал, которого надо много.
В этот раз рассмотрим, как формируется коллаген внутри клетки.
Начало синтеза
- В ядре в виде иРНК считывается информация с генов, которые кодируют альфа-цепи. От того, какие альфа-цепи сформируют итоговую молекулу, зависит тип коллагена.
- Затем иРНК (информационная РНК) выходит из ядра в цитоплазму и попадает в гранулярную или шероховатую эндоплазматическую сеть (шЭПС). На её рибосомах образуется цепь пре-про-полипептида.
- Пре-про-полипептид – это предшественник альфа-цепи. Он содержит большие глобулярные пропептиды на концах. Вы можете также встретить термины «про-альфа-цепи» (pro-alpha-chains) и «препроколлаген». Пропептиды на концах молекулы не дают ей сформировать спираль и объединиться с другими альфа-цепями раньше времени.
- В шЭПС происходит дальнейшая посттрансляционная модификация.
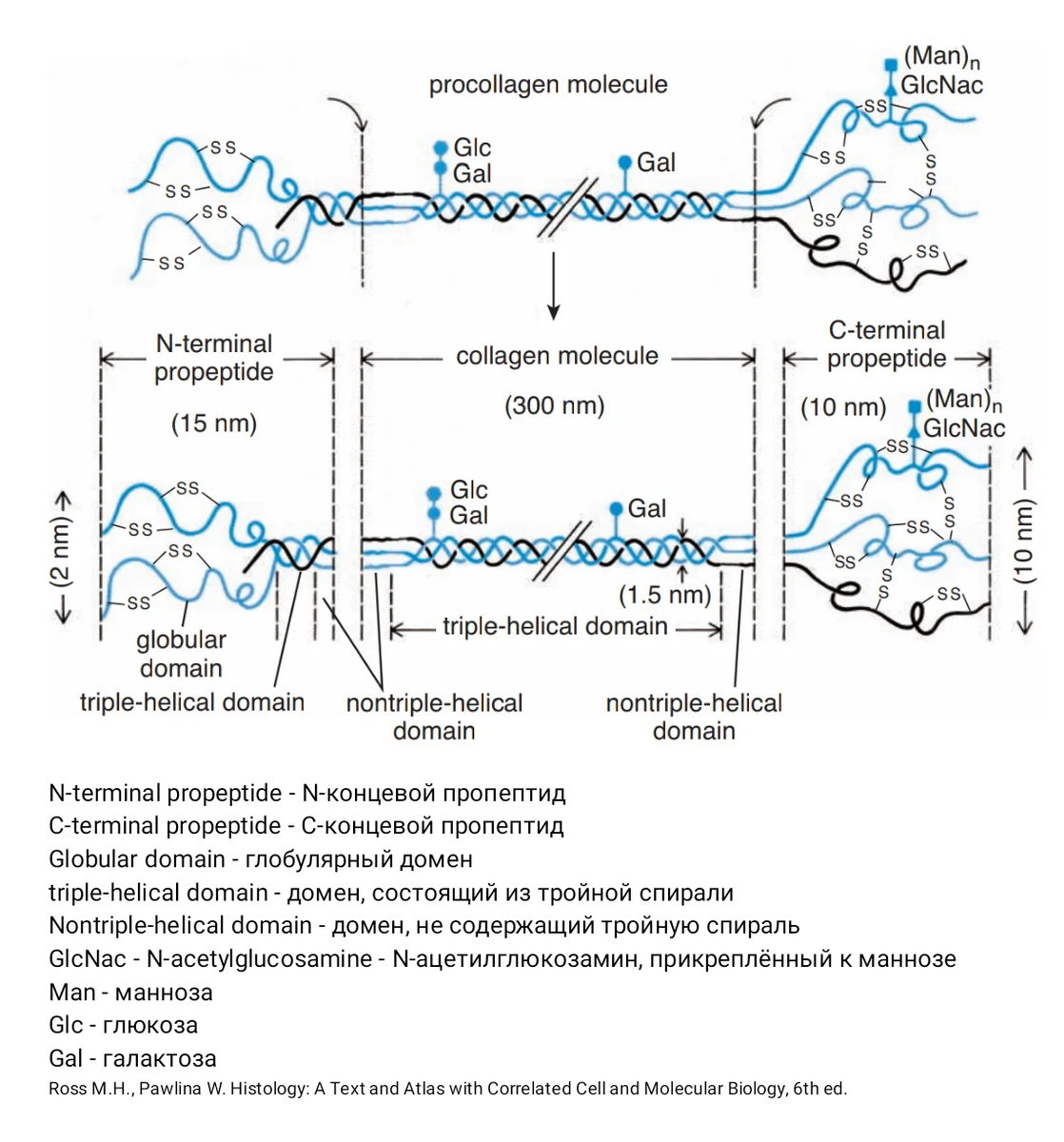
Посттрансляционная модификация
Пока всё шло стандартно: транскрипция информации с ДНК на иРНК, затем трансляция в шЭПС. Дальше интереснее: посттрансляционная модификация – это многостадийное превращение пре-про-полипептида в проколлаген. Итак, что происходит:
- Удаляется сигнальная последовательность на N-конце.
- К радикалам лизина и пролина присоединяются гидроксильные (OH-) группы. За это отвечают ферменты гидроксилазы (пролил 4-гидроксилаза, пролил 3-гидроксилаза, лизилгидроксилаза), кофактором которых является витамин C. Функция пролил 3-гидроксилазы ещё мало изучена, её инактивация приводит к тяжёлой форме несовершенного остеогенеза (osteogenesis imperfecta).
- Благодаря гидроксилированию остатки пролина и лизина формируют водородные связи для удержания пространственной структуры коллагена. После этого полипептид сможет принять форму спирали.
- Происходит гликозилирование ОН-групп лизина молекулами галактозы и глюкозы с помощью галактозилтрансфераз и глюкозилтрансфераз. Возможно, существует связь между гликозилированием коллагена и ревматоидным артритом, системной склеродермией, образованием злокачественных опухолей.
- Ещё такой момент. До формирования тройной спирали альфа-цепи должны быть стабилизированы дисульфидными мостиками внутри самой цепи.
Теперь три гидроксилированные и гликозилированные про-альфа-цепи собираются в тройную спираль, образуется молекула проколлагена. Концевые пептиды ещё не удалены. Сборка тройной спирали начинается с C-концевого домена и продолжается по направлению к N-концевому домену, образуя молекулу проколлагена.
Эта конструкция транспортируется в аппарат Гольджи для заключительной модификации. В аппарате Гольджи молекулы упаковываются в секреторные пузырьки, чтобы выйти во внеклеточное пространство.
Формирование тройной спирали и выход проколлагеновой молекулы из ЭПС зависит от шаперона HSP47. Он связывается с триплетами «глицин – X – аргинин» разных типов коллагенов. Мутации HSP47 также являются причиной несовершенного остеогенеза (osteogenesis imperfecta).