Семена служат отправной точкой вегетативного роста растения. В семенах происходят важные процессы развития, необходимые для дальнейшего роста и развития нового растения. Покой семян, одно из наименее изученных явлений, однако кое что уже известно. Давайте разбирать…
- Структура семян
Один из основных факторов покоя семян - их строение, ведь именно оно определяет как будет проникать влага в семя. Хотя семена двудольных и однодольных растений отличаются (в основном наличием эндосперма, хотя бывают и исключения), семенная оболочка присутствует у всех и именно она не даёт влаге проникать в глубь семени.
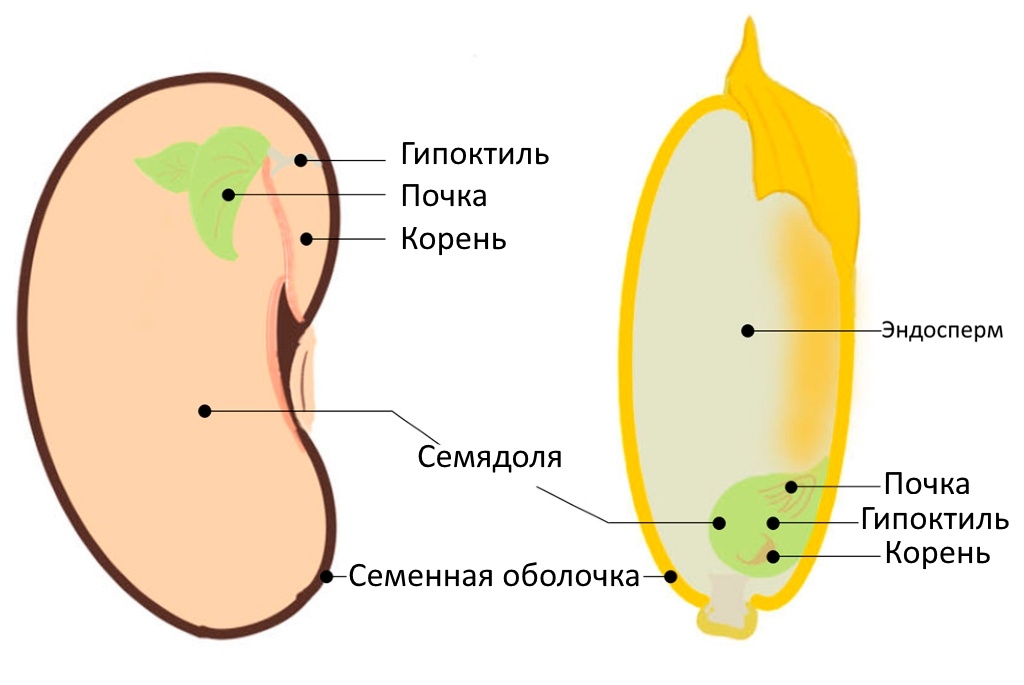
Точнее сказать не полностью блокирует, а значительно замедляет проникновение воды. Когда это происходит, то семя возобновляют свою метаболическую активность. Это позволяет им завершить необходимую клеточную деятельность при подготовке к последующему росту проростков.
При насыщении водой корешок продолжает удлиняться, пока не прорвет семенную кожуру и в конечном итоге не вырвется наружу. После прорыва корешка гипокотиль начинает удлиняться, а затем эмбрион растет. Выход корешка из семенной кожуры означает завершение процесса прорастания семени.
- Изменение питательных веществ
Семена потребляют собственные запасенные вещества, чтобы обеспечить энергию, необходимую для прорастания. В зрелых сухих семенах многие физиологические и биохимические процессы с участием ферментов не протекают из-за отсутствия подходящих условий или отсутствия ключевых компонентов.
Как только условия, необходимые для реакции, достигнуты, при совместном действии этих ферментов запасённые вещества, такие как крахмал, жир и белок, превращаются в сахар посредством окислительного дыхания, что делает их транспортабельными и пригодными для использования. На протяжении всего периода покоя семена не осуществляют физиологической деятельности, и реактивация роста в конце периода покоя требует притока энергии, которую как раз обеспечивает сахар.
2.1. Крахмал
Крахмал содержит амилозу и амилопектин, имеющие как линейные, так и разветвленные молекулы α-1,4-глюкозидного типа с α-1,6-связями в точках ветвления. Аденозин-5'-дифосфатглюкоза (АДФ-глюкоза), вещество-предшественник синтеза крахмала, синтезируется АДФ-глюкозопирофосфорилазой.
Крахмал обычно производится и откладывается в запасающих клетках, в семядолях или в эндосперме, служа долгосрочными источниками углерода для будущего роста.
Во время прорастания семян крахмал гидролизуется амилазой с образованием микромолекулярного декстрина, который далее гидролизуется с образованием мальтозы и. Конечный гидролиз осуществляет мальтаза с образованием глюкозы, которая и используется в качестве источника энергии.
В зависимости от типа изомеризации продукта ферментативного гидролиза эти амилазы можно разделить на α-амилазы, β-амилазы, γ-амилазы и амило-α-1,6-глюкозидаза.
Полученные сахара играют решающую роль в прорастании семян, поскольку они обеспечивают энергию за счет расщепления глюкозы посредством гликолиза и других путей метаболизма сахара.
2.2. Жир
Жир, основной энергетический запас семян масличных культур, содержится в семенах бобовых в виде триацилглицеринов. Во время начального созревания масличных культур вырабатываются большие количества свободных жирных кислот, которые в основном используются для синтеза жиров. Во время созревания семян происходит превращение насыщенных жирных кислот в ненасыщенные жирные кислоты.
Во время прорастания триацилглицерины разлагаются липазой на глицерин и свободные жирные кислоты, способствуя росту проростков. Эти свободные жирные кислоты впоследствии расщепляются и в конечном итоге превращаются в сахара.
Жир — это неполярное соединение, хранящееся в безводном состоянии. Обменная энергия жира в шесть раз превышает энергию гомополисахарида гликогена(который также хранится в семенах) той же массы.
Катаболизм жирных кислот происходит в митохондриальном матриксе, где жирные кислоты сначала активируются коферментом-А тиокиназой, что позволяет им проникнуть в митохондрии для β-окисления. Этот процесс приводит к образованию ацетил-КоА, который впоследствии генерирует энергию за счет цикла трикарбоновых кислот.
2.3. Белок
Белки играют незаменимую роль в основных функциях клеток и живых организмов. Во время прорастания сохраненные белки гидролизуются протеазами, высвобождая аминокислоты и низкомолекулярные пептиды.
Аминокислоты могут обеспечивать энергию посредством окисления углеродных цепочек после дезаминирования или служить строительными кирпичиками для синтеза новых белков.
Кроме того, аминокислоты также могут превращаться в промежуточные продукты метаболизма, такие как пируват, оксалоацетат и α-кетоглутарат. По сути, они являются «топливом» для различных обменных процессов внутри организма.
Модификации фосфорилирования белков также имеют решающее значение в регуляции покоя и прорастания семян. В настоящее время четыре следующих первичных киназы широко исследованы в состоянии покоя и прорастания семян: митоген-активируемые (MAPK), кальций-зависимые (CDPK), неферментируемые сахарозой (SnRK) и рецептороподобные протеинкиназы (RLK). ). Эти киназы в первую очередь регулируют покой и прорастание, реагируя на сигналы фитогормонов.
- Передача сигналов эндогенных гормонов и регуляция генов
Фитогормоны жизненно важны для регуляции покоя и прорастания семян. Согласно доктрине эндогенной гормональной регуляции, предложенной Ханом в 1975 году, взаимодействие гиббереллиновой кислоты (ГК), цитокинина (ЦТК) и абсцизовой кислоты (АБК) определяет покой и прорастание семян.
Например:
- Когда в семенах присутствуют и АБК, и ГК, эффект ГК, способствующий прорастанию, ингибируется АБК;
- При одновременном присутствии ГК, ЦТК и АБК ЦТК оказывает антиингибирующее действие, способствуя прорастанию;
Другие эндогенные гормоны, такие как этилен (ЭТН), брассинолид (БР), ауксин(АУ) и стриголактоны (СЛ), также участвуют в регуляции физиологических и биохимических процессов покоя и прорастания семян.
3.1. Абсцизовая кислота
АБК является важным фитогормоном, регулирующим период покоя. Индукция покоя во время развития семян тесно связана с повышением уровня эндогенной АБК в растениях.
У арабидопсиса существует четыре типа белков, связывающих АБК, а именно: локус контроля времени цветения A (FCA), рецептор ABA (ABAR), рецептор, связанный с G-белком (GPCR, 7-трансмембранный рецептор), и устойчивость к пирабактину (PYR) / PYR1-LIKE. (PYL)/РЕГУЛЯТОРНЫЙ КОМПОНЕНТ РЕЦЕПТОРА ABA (RCAR). Среди них FCA действует как РНК-связывающий белок, который контролирует время цветения. ABAR, подобно субъединице Mg-хелатазы H бобовых белков (ChlH), продуцирует Mg-протопорфирин Ⅸ (Mg-прото), который координирует экспрессию ядерных и хлоропластных генов. Эта координация, в свою очередь, вызывает реакцию нижестоящего компонента ABSCISIC ACID INSENSITIVE4 (ABI4).
Подробный механизм действия этого гормона на семена заслуживает отдельной статьи, так как хоро изучен. Если вам нужен это хардкор обязательно намекните.
3.2. Ауксин
Ауксины действуют как ингибитор прорастания семян у многих геофитов. В случае Arabidopsis ауксин играет ключевую роль как в обеспечении покоя, так и в ингибировании прорастания за счет усиления передачи сигналов ABA.
Молекулярные механизмы, лежащие в основе этих процессов:
В регуляции ауксина участвуют белки F-бокса, в частности белки TRANSPORT INHIBITOR RESPONSE1 (TIR1)/AUXIN SIGNALING F-BOX (AFB), белки ауксина/индол-3-уксусной кислоты (Aux/IAA) и фактор ответа на ауксин (ARF). При восприятии передачи сигналов ауксина с помощью TIR1/AFB Aux/IAA рекрутируется для деградации через протеасомный путь комплекса SCF TIR1/AFB -26S, который облегчает ингибирование транскрипционных факторов ARF, чтобы вызвать или заглушить экспрессию нижестоящих ауксинов. чувствительные гены .
3.3. Гиббереллиновые кислоты
ГК представляют собой многофункциональные физиологические регуляторы, участвующие в различных аспектах развития растений, таких как удлинение побегов, развитие корней, цветение и прорастание семян. Было замечено, что эндогенные ГК способствуют выпячиванию корешков, тем самым ускоряя прорастание семян. В семенах пшеницы и риса ГК являются основными фитогормонами, регулирующими период покоя, где баланс АБК/ГК имеет решающее значение для степени покоя, при этом более высокая концентрация ГК приводит к меньшему периоду покоя.
При восприятии ГК ядерным рецептором GIBBERELLIN-INSENSITIVE DWARF1 (GID1) комплекс GA-GID1 рекрутирует ингибиторы роста DELLA (DELLA включает GAI, RGA и RGL2), членов семейства GRAS, которые действуют как репрессоры роста растений. Впоследствии комплекс SCF SLEEPY1 (SLY1)/GID2 убиквитинирует DELLA, маркируя их деградацию в присутствии 26S протеасомы. Кроме того, на регуляцию ГК влияет температура. Например, SOMNUS (SOM) кодирует белок цинковых пальцев CCCH-типа. ГК дезактивирует СОВ и способствует прорастанию семян при низких температурах, однако при высоких температурах экспрессия SOM усиливается DELLA и ABA, а эпигенетически - комплексом AGAMOUS-LIKE 67-EARLY BOLTING IN SHORT DAYS (AGL67-EBS), ингибирующим синтез ГК и предотвращающим прорастание семян. При сверхвысоких температурах (выше 32 °C) взаимодействие между БЕЛКОМ ТЕПЛОВОГО ШОКА (HSP) и ФАКТОРОМ ТЕПЛОВОГО ШОКА (HSF) активирует синтез и накопление белка FUSCA3 (FUS3), запуская синтез АБК и деградацию ГК, тем самым препятствуя прорастанию семян.
3.4. Цитокинины
Эффект ЦTK растительный гормон, играющий решающую роль в прорастании семян. Но действие его очень видоспецифично активное чем ГК.
Открытие ЦTK как растительных гормонов со свойствами, способствующими делению клеток, датируется 1955 годом, когда Миллер и Скуг исследовали стимулирующее рост воздействие ДНК спермы сельди, полученной в автоклаве, на ткань табачного каллуса.
У растений путь ЦTK регулируется четырьмя чередующимися последовательными событиями фосфорилирования, которые включают His киназу, домен приемника Asp, белки-фосфопереносчики гистидина Arabidopsis (AHP) и отдельные регуляторы ответа (RR), где His киназа и Asp домен-приемник вместе образуют «гибридный» рецептор гистидинкиназы (НК).
Рецептор HK имеет консервативный цитокинин-связывающий экстрацитозольный домен CHASE (циклазы/гистидинкиназа-ассоциированный чувствительный внеклеточный домен), по меньшей мере два трансмембранных домена, цитозольную область, содержащую домен гистидинкиназы, канонический домен-приемник и дивергентный домен-приемник с маловероятным функциональность. Такое структурное расположение позволяет цитокинину связываться с доменом CHASE и активировать цитозольный домен гистон-киназы. Эта активация приводит к аутофосфорилированию консервативного остатка His с последующим переносом фосфатной группы на консервативный Asp внутри домена-приемника. Это событие фосфорилирования впоследствии передается нижестоящим белкам AHP и RR, создавая положительную регуляторную цепь. В конечном итоге сигнал цитокинина запускает транскрипционные изменения в ядре.
3.5. Этилен
ЭТН - газообразный фитогормон, в зависимости от дозы может менять период покоя семян и способствовать их прорастанию. Было обнаружено, что экзогенное применение ЭТН значительно способствует прорастанию семян хлопчатника и Fraxinus mandshurica в условиях осмотического стресса, что позволяет предположить, что ЭТН может действовать как положительный регулятор, высвобождая семена из состояния покоя.
Напротив, было обнаружено и то, что повышенные концентрации ЭTH не только задерживают прорастание семян, но и подавляют рост корешков семян перца. Это ингибирование прорастания семян с помощью ЭTH также наблюдалось в семенах Camellia Oleifera.
Биосинтез ETH следует хорошо изученному пути. Первоначально метионин (Met) катализируется метионин-аденозилтрансферазой (MAT) с образованием S-аденозилметионина (SAM); Затем SAM превращается в 1-аминоциклопропан-1-карбоновую кислоту (АСС) посредством АСС-синтазы (ACS). Последующее окисление АСС оксидазой АСС (ACO) приводит к образованию ETH, включающему круговую реакцию, известную как цикл Янга. У арабидопсиса были идентифицированы пять мембранно-локализованных рецепторов ЭTH: этиленовый ответ 1 (ETR1), этиленовый ответ 2 (ETR2), устойчивый к этилену 1 (ERS1), устойчивый к этилену 2 (ERS2) и нечувствительный к этилену 4 (EIN4). Связывание ЭTH с его рецепторами, такими как ETR1, деактивирует конститутивную протеинкиназу тройного ответа 1 (CTR1) через домен HK ETR1. И наоборот, этот процесс активирует киназный каскад, который регулирует нечувствительный к этилену 2 (EIN2) и его внутриядерные факторы транскрипции, включая нечувствительный к этилену 3 (EIN3), EIN3-LIKE (EIL) и белки, связывающие элементы ответа на этилен (EREBP) / факторы, чувствительные к этилену. (ERFs), что в конечном итоге приводит к транскрипции генов ответа на этилен.