
Воспользовавшись тем что пищеварительная система хрюшек схожа с человеческой, ученые поставили на них ряд опытов и получили ответы о том как хлеб из однозернянки в отличии от хлеба из обычной пшеницы влияет на уровень глюкозы и инсулина, а также состав микробиоты кишечника.
Эффекты In Vivo хлеба из однозернянки (Triticum monococcum) на микробиоту кишечника, метаболом и гликемические и инсулинемические реакции у свиней
Аннотация: Пшеница однозернянка (Triticum monococcum) характеризуется высоким содержанием белков, биологически активных соединений, таких как полиненасыщенные жирные кислоты, фруктаны, токолы, каротиноиды, алкилрезорцины и фитостеролы, и более низкой активностью α-, β-амилазы и липоксигеназы по сравнению с полиплоидной пшеницей. Эти особенности делают муку из однозернянки хорошим выбором для приготовления более здоровой пищи. В настоящем исследовании мы исследовали влияние хлеба из однозернянки (EB) на физиологию и метаболизм кишечника свиней, характеризуя гликемические и инсулинемические реакции, а также профили микробиоты и метаболома. В исследовании приняли участие 16 товарных гибридных свиней: 4 свиньи были использованы для оценки постпрандиальных гликемических и инсулинемических реакций, а 12 свиней подверглись 30-дневному диетическому вмешательству для оценки изменений микробиоты и метаболизма после употребления EB или стандартного пшеничного хлеба (WB). Постпрандиальное повышение уровня инсулина после приема EB характеризовалось более низким абсолютным уровнем и, как также наблюдалось в отношении глюкозы, двухфазным по сравнению с таковым в ответ на прием WB. Потребление EB привело к обогащению кишечной микробиоты продуцентами короткоцепочечных жирных кислот (e.g., Blautia, Faecalibacterium, и Oscillospira) и к более высокому метаболическому разнообразию при меньшем содержании сукцината, что, вероятно, связано с улучшением всасывания и, следовательно, способствует глюконеогенезу в кишечнике. Наблюдаемые изменения, как в структурном, так и в метаболическом отношении, убедительно свидетельствуют о том, что потребление EB может поддерживать благоприятную для здоровья конфигурацию экосистемы кишечника.
Ключевые слова: однозернянка; кишечная микробиота; кишечные метаболиты; гликемическая реакция; инсулинемическая реакция
Введение
Пшеница однозернянка (Triticum monococcum) была одной из первых культур, одомашненных примерно 12 000 лет назад на Ближнем Востоке, наряду с полбой (Triticum dicoccum). Как правило, однозернянка выращивалась на малоплодородных сельскохозяйственных землях, так как мог выживать в суровых условиях и на бедных почвах, где другие виды пшеницы были не способны выжить. Спельта (Triticum spelta) представляет собой гексаплоидную серию генома Triticum, которая характеризуется отличной адаптацией к широкому спектру условий окружающей среды [1]. По сравнению с полиплоидной пшеницей, она отличается более высоким содержанием белков и некоторых хорошо известных биологически активных соединений, таких как полиненасыщенные жирные кислоты, фруктаны, токолы, каротиноиды, алкилрезорцины, фитостеролы, а также более низкой активностью α-, β-амилазы и липоксигеназы [2]. Эти особенности состава делают муку из однозернянки хорошим выбором для приготовления более полезных продуктов. В частности, предполагается, что наличие антиоксидантных соединений и белковый состав имеют некоторую взаимосвязанность с уменьшением сердечно-сосудистых заболеваний и гипоаллергенностью, соответственно. В частности, было показано, что однозернянка экспрессирует небольшое количество стимулирующих T-клетки глютеновых пептидов, что имеет важное значение при развитии целиакии [3]. Ферментированный в лабораторных условиях хлеб из однозернянки продемонстрировал более высокий уровень каротиноидов по сравнению с современными пшеничными хлебами и более выраженный противовоспалительный эффект в клетках кишечного эпителия Caco-2 [4]. Учитывая решающую роль кишечной микробиоты в метаболизме пищевых соединений, включая биоактивацию растительных полифенолов в полезные для здоровья метаболиты и производство короткоцепочечных жирных кислот (SCFA, в основном ацетата, пропионата и бутирата) в результате ферментации клетчатки, которые являются основными регуляторами физиологии организма-хозяина [5], также разумно предположить аналогичную пользу однозернянки для здоровья in vivo. Однако, несмотря на то, что в течение последних нескольких лет велись ожесточенные споры о том, насколько полезны для здоровья человека древние сорта зерновых, выращиваемые по всему миру, на сегодняшний день лишь в нескольких исследованиях изучалось их воздействие in vivo [1,6]. В частности, для однозернянки, одного из наиболее репрезентативных древних злаков, результаты исследований in vitro показали хороший потенциал для здоровья из-за его влияния на концентрацию глюкозы и инсулина в крови, что позволяет использовать продукты на основе однозернянки при метаболических заболеваниях [7,8], но никто не рассматривал изменения в структуре микробиоты, а также в наборе метаболитов в кишечнике, потенциально влияющих на множество метаболических и иммунологических путей, которые имеют отношение к здоровью хозяина. В попытке восполнить этот пробел, мы исследовали микробиоту и метаболизм кишечника свиней, которых кормили хлебом из однозернянки, в сравнении с пшеничным хлебом. Свиньи имеют значительное анатомическое и физиологическое сходство с людьми, особенно в том, что касается строения кишечника, с сопоставимым временем прохождения и аналогичными процессами пищеварения и всасывания [9,10]. Более того, как и люди, они по-настоящему всеядны, в отличие от других потенциальных моделей млекопитающих, таких как собаки, кошки, жвачные животные, кролики и грызуны, которые эволюционно выработали альтернативные стратегии пищеварения. Наконец, и свиньи, и люди являются ферментерами толстой кишки и имеют схожий состав микробиоты толстой кишки. Все эти особенности свиней делают их незаменимыми для проведения опытов в области питания человека [11,12]. В настоящем исследовании на модели свиньи мы исследовали влияние 30-дневного употребления хлеба из однозернянки и пшеничного хлеба на экосистему кишечника с помощью секвенирования гена 16S rRNA нового поколения и метаболомики образцов фекалий, а также образцов из отделов подвздошной кишки и толстой кишки. Была проведена оценка последствий употребления однозернянки и пшеничного хлеба на физиологию животных, показатели крови, гликемию после приема пищи и инсулина.
Материалы и методы
Экспериментальные сорта хлеба
Экспериментальный хлеб, который использовался в данном исследовании, был приготовлен пекарской компанией Vini, Польша. Два вида хлеба были изготовлены из обычной белой пшеничной муки и муки из однозернянки, соответственно. Их замешивали на пекарских дрожжах (2,5% муки) при температуре 30 °C в течение 1,5 ч, а затем выпекали при температуре 195 °C в течение 45 мин. для получения стандартного пшеничного хлеба и хлеба из однозернянки (WB и EB, соответственно). WB и EBбыли высушены и измельчены, а затем доставлены на факультет ветеринарной медицины Болонского университета для кормления экспериментальных животных.
Описание животных и исследования
Все мероприятия, проводимые в рамках этого исследования, регламентировались протоколами, которые были одобрены Министерством здравоохранения Италии в соответствии с D.Lgs n.116/92, а также соответствовали местным руководствам по использованию лабораторных животных. Исследование было разделено на два эксперимента: эксперимент 1 (Tr1) целью которого была оценка гликемической и инсулиновой реакции после приема WB и EB; и, эксперимент 2 (Tr2) целью которого была оценка кишечной и фекальной микробиоты и состава метаболома после 30-дневной WB или EB диеты. Животные, приобретенные на местной ферме, были доставлены на факультет ветеринарной медицины Болонского университета за 5 дней (период акклиматизации) до начала экспериментов. В течение периода акклиматизации всех свиней кормили стандартным рационом для свиней (SD). Стандартный рацион был специально разработан для свиней весом от 30 до 80 кг компанией CESAC s.c.a. Conselice (Ravenna, Italy). Состав рациона приведен в дополнительной таблице S1.
В эксперименте Tr1, 4м товарным гибридным свиньям Landrace x Duroc (самки весом 50 ± 2 кг, возраст - 14 недель) хирургическим путем под общим наркозом ввели в яремную вену центральный постоянный силиконовый катетер (BBraun, Bethlehem, PA, USA) и поместили в отдельные подвесные боксы. Перед операцией свиньям предварительно вводили внутримышечно (в/М (IM)) болюс кетамина (20 мг/кг) и медетомидин (0,03 мг/кг). Через 10 минут после этого, в положении лежа, был обеспечен венозный доступ через заднюю ушную вену, и животным вводили тиопентал болюсно (0,25 г на животное). Этот же венозный доступ использовался для инфузионной терапии (Lacteted Ringer, 10 mL/kg/h) и был удален в конце хирургической процедуры. Оротрахеальная интубация проводилась с использованием эндотрахеальной трубки с внутренним диаметром (ID) 8 мм. Анестезию поддерживали с помощью изофлурана (2-3%) в смеси кислорода и воздуха в соотношении 1:1. Через неделю после выздоровления животных приучили съедать 50 г экспериментального хлеба в течение 5 минут. Тренировки проводились в 8 часов утра, после 15-часового голодания в течение 7 дней подряд. В конце периода обучения (7 дней), утром, после 15-часового голодания, свиньям скармливали 50 г экспериментального хлеба или 50 г глюкозы (жидкого раствора) в качестве контрольного корма (С). Каждому животному случайным образом назначали диету (WB, EB или C) на девять дней, чтобы повторить каждую диету три раза. Образцы крови брали из яремной вены за 15 мин до введения экспериментального хлеба и каждые 15 мин в течение 2 ч для измерения уровня глюкозы и инсулина.
В эксперименте Tr2, 12 свиней (6 кастрированных самцов M и 6 самок F весом 36 ± 4 кг, 10-недельного возраста) были случайным образом разделены на четыре группы (по три животных в каждой группе) в соответствии с их полом. В течение 30 дней животных кормили два раза в день смесью экспериментального хлеба и SSD в соотношении 1:1, 2 группы (n = 6; 3 M и 3 F) ели WB и две группы (n = 6; 3 M и 3 F) ели EB. Животные получали по 1 кг смешанного рациона, разделенного на 2 раздельных приема пищи, воду давали в неограниченном количестве. Животных взвешивали на 0, 7, 14 и 30 день; для классификации кала каждой ячейке ежедневно присваивали оценку: 0 для нормального кала; 0,5 для пушистого/кашицеобразного кала; и 1 балл для водянистых выделений. У каждого животного отбирали образцы кала, как описано в разделе "Отбор проб", через 0, 15 и 30 дней для оценки микробиоты кишечника и изучения различий в процессах пищеварения с помощью методов NMR (Метод ядерного магнитного резонанса (ЯМР)) и газовой хроматографии-масс-спектрометрии (ГХ-МС (gas chromatography-mass spectrometry (GC-MS)). Образцы крови (в пробирках K3-EDTA) были взяты в начале и в конце исследования для клинической оценки (общий анализ крови – CBC).
Взятие проб и анализ крови
В случае Tr1 кровь, взятую из яремного катетера, немедленно анализировали с помощью экспресс-глюкометра (Contour Glucometer Xt, Bayer S.p.a., Leverkusen, Germany) для определения уровня глюкозы в крови, в то время как кровь для количественного определения инсулина собирали в пробирку K3EDTA и хранили при температуре +4 °C до окончания периода отбора проб после приема пищи. Затем пробирки с K3EDTA центрифугировали (1000 г × 10 мин) и плазму хранили при температуре -20 °C. Уровень инсулина определяли в плазме крови с помощью хемилюминесцентного иммуноанализа в Veterinary Clinical Pathology Service of DIMEVET (UNIBO). Сначала рассчитывали площадь под кривой (AUC), а затем гликемический и инсулинемический индексы (GI и II) для каждого рациона в следующем соотношении [GI и II = AUC продукта/среднее значение (AUC контрольная) × 100] [13]. Диетическая гликемическая и инсулинемическая нагрузка (GL и IL) рассчитывалась с учетом количества углеводов в каждом хлебе [GL и IL = (GI или II × г. доступных углеводов)/100] [14]. В случае Tr2, образцы крови в начале и в конце исследования были взяты из яремной вены под общей анестезией по следующему протоколу: внутримышечное (IM) введение азаперона (2 мг/кг), через 10 мин в темноте и в спокойном состоянии внутримышечная инъекция кетамина (40 мг/кг). Пластиковые пробирки VenosafeQR для K3EDTA (Terumo, Tokyo, Japan) были использованы для взятия крови, бразцы были проанализированы в Veterinary Clinical Pathology Service of DIMEVET (UNIBO) для общего анализа крови (CBC). Для отбора проб кала оператор находился в боксе с животными. Образцы отбирали непосредственно при спонтанной дефекации перед загрязнением и сразу же хранили при температуре -20 °C в течение 24 часов. Затем образцы хранили при температуре -80 °C до момента проведения анализа. В конце исследования животных усыпляли с помощью 0,3 мл/кг TanaxQR (MSD Animal Health Italia, Milano, Italy) под общим наркозом для взятия проб содержимого желудочно-кишечного тракта и слизистых оболочек (толстой и тонкой кишки). Вкратце, пищеварительный тракт был удален и разделен на анатомические части, было выполнено выскабливание слизистой оболочки подвздошной кишки и толстой кишки, как описано ранее [12].
Извлечение микробной ДНК, секвенирование Illumina MiSeq и обработка данных
Общая бактериальная ДНК была выделена из кала и соскобов со слизистой оболочки подвздошной и ободочной кишок с использованием многократного измельчения бусинами вкупе с колоночной хроматографией [15], с небольшими изменениями [16]. Вкратце, около 250 мг каждого образца суспендировали в 1 мл буфера для лизиса (500 mM NaCl, 50 mM Tris-HCl pH 8, 50 mM EDTA, 4% SDS) и трижды взбивали с помощью прибора FastPrep-24 Instrument (MP Biomedicals, Irvine, CA, USA) при 5,5 движениях в секунду в течение 1 минуты при наличии 4х стеклянных шариков диаметром 3 мм и 0,5 г циркониевых шариков диаметром 0,1 мм BioSpec Products, Bartlesville, OK, USA). После 15-минутной инкубации при 95°C и центрифугирования на полной скорости в течение 5 мин для гранулирования сырья нуклеиновые кислоты осаждали путем добавления 10 М ацетата аммония и одного объема изопропанола с последующей инкубацией во льду. Гранулы промывали 70%-ным этанолом, ресуспендировали в TE-буфере и обрабатывали 10 мг/мл DNase-free RNase при 37 °C в течение 15 минут. Обработку протеиназой К и очистку ДНК на колонках проводили в соответствии с инструкциями производителя (QIAamp DNA Stool Mini Kit; QIAGEN, Hilden, Germany). Парное секвенирование гипервариабельной области V3-V4 гена 16S rRNA было выполнено на платформе Illumina MiSeq, в компании IGA Technology Services (http://igatechnology.com/; Udine, Italy). Считанные последовательности были сохранены в базе данных MG-RAST (https://www.mg-rast.org/linkin.cgi? project=mgp87784). Исходные последовательности были обработаны с использованием конвейера, объединяющего парный ассемблер последовательностей (PANDAseq) [17] и количественный анализ микробной экологии (QIIME) [18]. Высококачественные результаты считывания были сгруппированы в оперативные таксономические единицы (OTUs) с 97%-ным сходством с использованием UCLUST [19]. Таксономия была определена с использованием классификатора Ribosomal Database Project (RDP) в соответствии с базой данных Greengenes (выпуск от мая 2013 г.). Химерные и одноэлементные OTU были отброшены. Альфа-разрежение было выполнено с использованием филогенетического разнообразия Faith, наблюдаемого OTU и индекса Шеннона. Бета-разнообразие было оценено путем вычисления взвешенных и невзвешенных расстояний UniFrac.
Метаболомика
Для описания метаболического профиля образцов кала были использованы два различных аналитических подхода, а именно протонный ядерно-магнитный резонанс (1H-NMR) ля жидкой фракции и газовая хроматография-масс-спектрометрия (GC-MS) для летучих соединений. 1H-NMR анализ был затем также распространен на содержимое подвздошной и ободочной кишок.
Образцы фекалий и образцы из кала и толстой кишки были подготовлены для анализа методом 1H-NMR по методике Bryszewska и соавт. [12]. Чтобы применить NMR в качестве количественного метода [20], задержка рециркуляции была установлена равной 5 с, при этом учитывалось время продольной релаксации исследуемых протонов. Сигналы были определены путем сравнения их химического сдвига и множественности с помощью банка данных программного обеспечения Chenomx (Chenomx Inc., Edmonton, AB, Canada, ver 8.1) и литературных источников [21]. Исходный уровень спектров1H-NMR был скорректирован в соответствии с описанием Bryszewska и соавт. [12]. Различия в содержании растворимых твердых веществ между образцами учитывались путем нормализации вероятностных коэффициентов [22].
Для выявления летучих метаболитов в кале использовали твердофазный микроэкстракционно-газохроматографический и масс–спектрометрический анализ (SPME–GC–MS). Отбор проб, условия GC-MS и обработка данных проводились в соответствии с Taneyo Saa и соавт. [23]. Частичный дискриминантный анализ методом наименьших квадратов (PLS-DA) был использован для уменьшения размерности данных, а графики первых 2-х переменных позволили визуализировать временное разделение в каждой группе в соответствии с Dejean и соавт. [24]. Метод случайного леса - Random forest (RF) использовался в качестве метода отбора признаков для идентификации молекул, которые отвечают за различение времен (t0 and t30) в рамках каждой диеты и диет после вмешательства (t30). Выбор RF-признака был получен путем вычисления Importance Scores, т.е. среднего снижения точности, о чем сообщили Enot и соавт. [25]. Были рассчитаны коэффициенты корреляции Пирсона между 10 наиболее важными молекулами, идентифицированными с помощью RF. Переменные с коэффициентом корреляции более 0,7 рассматривались как относящиеся к группе, которая указывает на различные продукты распада во время диеты.
Статистика
Данные анализов крови и массы тела по формам Tr1 и Tr2 были проанализированы с помощью MedCalc Statistical Software version 16.4.3 (MedCalc Software bvba, Ostend, Belgium; https://www.medcalc.org; 2016). В частности, в Tr1 для измерения уровня глюкозы и инсулина данные были нормализованы и проанализированы с помощью непараметрического теста Kruskal–Wallis. Когда тест был положительным (p < 0.05), был проведен post-hoc анализ в соответствии с Conover, 1999 [26]. В Tr2 данные о CBCанализировались с помощью Wilcoxon test для парных образцов, в то время как данные о массе тела анализировались с помощью ANOVA test для повторных измерений. Для микробиоты и метаболома все статистические анализы, включая анализ главных координат (PCoA), ANOVA и непараметрические тесты (Wilcoxon test для парных или непарных данных, по мере необходимости), были выполнены в R (http://www.R-project.org). Значения p были скорректированы для множественных сравнений с использованием метода Benjamini–Hochberg. Скорректированное значение p < 0.05 считалось статистически значимым, если не указано иное. Что касается водорастворимого метаболома, то различия между двумя группами образцов были проанализированы с помощью непараметрического теста Wilcoxon, в то время как различия, вызванные употреблением обычной пшеницы и однозернянки при t30 в «подвздошная кишка - ободочная кишка – кал» в динамике были выявлены с помощью ANOVA-теста для повторных измерений. Надежные модели анализа главных компонент (PCA) были рассчитаны в соответствии с Hubert и соавт. [27]. Для образцов кала это было сделано на основе различий t30–t0, чтобы учесть парный характер данных.
Доступность данных
Наборы данных, сгенерированные и проанализированные в ходе текущего исследования, доступны соответствующим авторам по обоснованному запросу.
Результаты
Постпрандиальная гликемическая и инсулинемическая реакция
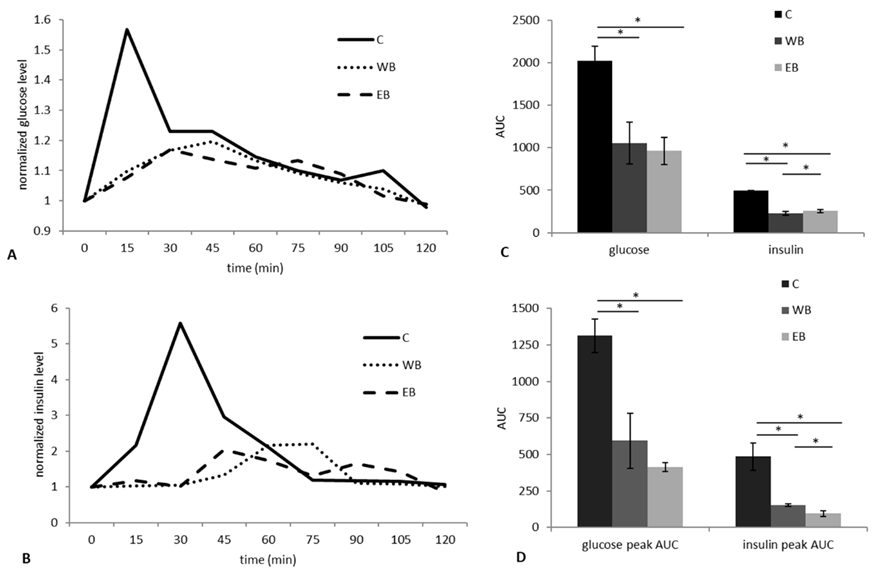
В первом эксперименте (Tr1) мы определили уровни глюкозы и инсулина в крови после приема пищи после употребления EB или WB в сравнении с эталонным продуктом (глюкозой) путем взятия крови из яремной вены свиньи в течение 2 часов после приема (Дополнительная таблица S2). Уровни глюкозы после приема пищи различались между WB и EB а 15–й и 105-й минутах, причем показатели EB были ниже (значение p = 0.0072 и p = 0.012, соответственно; Kruskal–Wallis test). На 15–й минуте постпрандиальный уровень инсулина после употребления WB был несколько выше (p = 0.012; Kruskal–Wallis test), чем после EB. Динамика содержания глюкозы и инсулина представлена на Рис. 1A,B. Форма волн гликемического и инсулинемического ответа различалась между WB и EB, будучи двухфазной у EB как для глюкозы, так и для инсулина. Для каждого экспериментального вида хлеба, а также для эталонного продукта были рассчитаны площадь под кривой (AUC), гликемический (GI) и инсулинемический (II) индексы, а также гликемическая (GL) и инсулинемическая (IL) нагрузка (Рис. 1C). GI составил 52.1 ± 12.1 и 47.5 ± 7.7 для WB и EB, соответственно. GL рассчитывали с учетом процентного содержания углеводов в экспериментальном хлебе (WB = 60.4%; EB = 59.9%) и она составила 31.5 ± 7.3 и 28.4 ± 4.6 для WB и EB, соответственно. Не было обнаружено различий между двумя диетами по показателям GI и GL, но обе отличались от контрольного продукта (p = 0.018; Kruskal-Wallis test). II и IL составили 46.2 ± 4.3 и 27.9 ± 2.6 для WB, и 51.7 ± 3.3 и 30.9 ± 2 для EB. Потребление EB привело к тому, что уровень II и IL после приема пищи был выше, чем у WB (p = 0.01; Kruskal–Wallis test). Согласно анализу пиков, AUC первых пиков глюкозы и инсулина, которые были получены при EB, были ниже, чем при WB, но разница была статистически значимой только для инсулина (p = 0.024; Kruskal-Wallis test) (Рис. 1D).
Общий анализ крови (CBC), количества кала и масса тела
Во втором эксперименте (Tr2), мы исследовали влияние 30-дневного употребления EB или WB на физиологию животных и экосистему кишечника. Данные общего анализа крови (CBC) и массы тела приведены в дополнительной таблице S3 и на дополнительном рисунке S1. Существенных различий в приросте массы тела между двумя диетами через 7, 15 и 30 дней не наблюдалось. Масса тела увеличилась на обеих диетах с t0 до t15 (p < 0.01; ANOVA для повторных измерений) и достигла фазы плато с t15 до конца исследования. Что касается CBC, то уровни гематокрита, гемоглобина, красных кровяных телец (эритроцитов) и ретикулоцитов увеличились в ходе исследования с t0 до t30 в обеих группах, что соответствовало физиологическим референтным интервалам для данного вида свиней [28]. Показатели воспалительных процессов и иммунологические показатели крови не изменились несмотря на то, что в первые 10 дней исследования у обеих групп наблюдалась умеренная диарея, которая, вероятно, была вызвана изменением рациона питания (дополнительная таблица S4). В частности, в течение первой недели кал был водянистым, а на второй неделе стал кашеобразным (пушистым). Затем был рассчитан балл, основанный на консистенции стула. Согласно распределению показателей кала, водянистый кал больше не выявлялся после t7 и t15 в группе EB и WB соответственно. Во второй части исследования (t15–t30) в обеих диетических группах фекалии всех свиней были в норме.
Реакция кишечной микробиоты на вмешательство в процесс питания
Образцы кала, собранные на 0, 15 и 30-й дни вмешательства в процесс питания (Tr2) были подвергнуты секвенированию гена 16S rRNA. Было получено и проанализировано в общей сложности 351 786 высококачественных считываний (среднее значение по выборке, 9772; диапазон, 7207–11,734). Считывания были сгруппированы в 16,236 OTUs с 97%-ным сходством. Согласно Shannon index, альфа-разнообразие значительно увеличилось после 30 дней рациона в обеих группах свиней, независимо от вида употребляемого хлеба (t0 против t30, WB, 7.07 против 7.58; EB, 6.93 против 7.58; p ≤ 0.04, Wilcoxon test). Аналогичным образом, взвешенное и невзвешенное распределение расстояний UniFrac показало разделение между образцами t0 и t30 как для группы EB, так и для группы WB (p ≤ 0.003, тест на перестановку с использованием псевдо-коэффициентов F) (Рисунок 2A,B). На типовом уровне в микробиоте фекалий свиней преобладали почти исключительно Firmicutes(средняя относительная численность при t0 составила 76,24%) и Bacteroidetes (18,48%), при этом Actinobacteria (1.67%) и Proteobacteria (1.68%) были субдоминантными компонентами (Рисунок 2C). Интересно, что обе диеты привели к одинаковому увеличению доли Spirochaetes, Lentisphaerae, и Tenericutes (p ≤ 0.04, Wilcoxon test), но только в группе EB наблюдалось значительное изменение доминирующей фракции микробиоты с увеличением представленности Bacteroidetes и соответствующим уменьшением Firmicutes (соотношение Firmicutes к Bacteroidetes: t0 против t30, 5.58 против 3.25; p = 0.04) (Рисунок 2C). Стоит отметить, что это изменение было заметно уже при t15 (соотношение Firmicutes к Bacteroidetes, 2.58; p = 0.002) (Дополнительный рисунок S2). С другой стороны, только потребление WB ривело к увеличению относительной численности Verrucomicrobia (p = 0.01) (Рисунок 2C). Таксономические сравнения на уровне рода подтвердили существование как общих, так и уникальных признаков в ответ на вмешательство в процесс питания (p < 0.05) (Рисунок 2D). Среди общих особенностей стоит отметить, что обе диеты, уже при t15, привели к увеличению численности Treponema, видов Ruminococcaceae (включая R. bromii), и бифидобактерий, и уменьшению доли представителей семейства Coriobacteriaceae, Acidaminococcus, и Megasphaera. Интересно, что только потребление EB способствовало обогащению следующих второстепенных компонентов: Oscillospira, Anaerovibrio, Paludibacter, неклассифицированных представителей отряда GMD14H09 (класс Deltaproteobacteria), и Blautia producta. И наоборот, количество бактерий Blautia spp. w было значительно снижено в микробиоте фекалий свиней, получавших WB, а также других производителей SCFA, таких как Faecalibacterium и Dorea formicigenerans. Потребление ВБ также привело к сокращению численности Collinsella, Streptococcus, Veillonellaceae (Mitsuokella и неизвестных родов), и Erysipelotrichaceae (Bulleidia и [Eubacterium]). С другой стороны, это способствовало обогащению неклассифицированных представителей отрядов Bacteroidales и Clostridiales. И, наконец, что еще более важно, после 30-дневного вмешательства в процесс питания микробиота фекалий свиней, которых кормили EB, отличалась от таковой у свиней, которых кормили WB, уменьшением относительного количества представителей семейства Bacteroidales S24-7 (EB против WB, 3.24% против 5.47%; p = 0.04).
Пристеночная микробиота, из отделов подвздошной кишки и толстой кишки
Соскобы слизистой оболочки из подвздошной и ободочной кишки были собраны после проведения диетологического вмешательства и подвергнуты секвенированию следующего поколения на основе гена 16S rRNA, как описано выше для образцов кала. Было получено в общей сложности 158 871 высококачественных считываний (среднее значение по выборке, 6620; диапазон, 3315–9913) которые были сгруппированы в 10,560 OTUs с 97% сходством. Согласно наблюдаемым показателям OTU, пристеночная микробиота после 30 дней употребления EB была более разнообразной, чем после введения WB (p = 0.02, Wilcoxon test) (Рисунок 3A). Аналогичным образом, только на уровне подвздошной кишки и только во взвешенном пространстве для определения расстояния, основанном на UniFrac, микробные фракции слизистой оболочки группировались в отдельные группы в зависимости от рациона питания (p = 0.04, тест на перестановку с использованием псевдо-коэффициентов F) (Рисунок 3B). В отличие от кала, в структуре микробиоты слизистой оболочки подвздошной кишки на уровне типа преобладали Firmicutes (реднее относительное количество, 39.28%) и Proteobacteria (26.77%), пропорции которых были противоположными и еще более дифференцированными на уровне слизистой оболочки толстой кишки (Firmicutes, 16.80%; Proteobacteria, 61.55%) (Рисунок 3C). В обоих отделах кишечника Bacteroidetes были основным субдоминантным типом, за ними следовали Chlamydiae в подвздошной кишке и Spirochaetes в толстой кишке. Интересно, что по сравнению со свиньями, питавшимися WB, фракция микробиоты, связанная со слизистой оболочкой подвздошной кишки, у свиней, получавших EB, была в значительной степени обогащена неклассифицированными микробами (EB против WB, 42.49% против 15.58%; p = 0.02, Wilcoxon test), в то время как в группе Firmicutes наблюдается значительное снижение (24.37% против 54.20%; p = 0.04) (Рисунок 3C). На уровне рода это различие было обусловлено, главным образом, недостаточной представленностью Streptoccoccus (в основном, вида S. alactolyticus) и Lactobacillus (в основном L. ruminis) в группе EB по сравнению с WB. Стоит отметить, что, хотя вид Bifidobacterium adolescentis и был в значительной степени субдоминантным, он был обнаружен во фракции слизистой оболочки подвздошной кишки только после потребления EB. На уровне слизистой оболочки толстой кишки потребление EB привело к снижению численности неизвестных видов семейства Ruminococcaceae, по сравнению с диетой, основанной на WB (Рисунок 3D).
Метаболомика водорастворимой фракции
Чтобы оценить влияние двух протестированных диет с точки зрения метаболомики, водорастворимые фракции содержимого подвздошной и ободочной кишок, а также кал были исследованы методом протонного ядерного магнитного резонанса (1H-NMR). Во-первых, мы уделили особое внимание глюкозе, SCFAs (то есть ацетату, пропионату и бутирату), сукцинату и лейцину, концентрация которых, как известно, строго связана с чувствительностью к инсулину [29–33] (Figure 4). Концентрация глюкозы, сукцината и лейцина в желудочно-кишечном тракте снижалась при обеих диетах. В рамках такой тенденции к снижению, диета с WB систематически приводила к повышению концентрации глюкозы и незначительному снижению концентрации сукцината в каждый момент времени по сравнению с диетой с EB. При обеих диетах наблюдалось заметное увеличение содержания SCFAs в ЖКТ, причем диета с WB ассоциировалась с более высоким содержанием ацетата и пропионата по сравнению с диетой с EB. В качестве второго шага в исследовании метаболомики мы расширили наши представления, применив 1H-NMR для нецелевого подхода к метаболомике. Можно было определить количество 70ти молекул, относящихся к химическим группам мономерных углеводов, аминокислот, SCFAs и органических кислот. Можно было количественно определить еще 15 молекул с частично неясной структурой, которые, следовательно, были обозначены местоположением в спектре NMR, выраженным в миллионных долях. Чтобы продемонстрировать специфические эффекты двух диет, мы сначала сосредоточились на фекалиях, рассчитав колебания t0–t30 для каждой молекулы и выбрав те, которые демонстрируют различия между двумя диетами с достоверностью выше 90% (Дополнительная таблица S5). Выделенные таким образом 5 молекул послужили основой для модели RPCA. На соответствующем графике оценки (Рисунок 5A), показатель PC 1, представляющий 98.7% от общей дисперсии между выборками, суммирует вариации t0–t30, при этом выборки, относящиеся к группе EB, в t30 характеризуются наивысшими оценками, а выборки, относящиеся к группе WB, в t30 характеризуются оценками, которые очень близки к таковым у образцов при t0. Таким образом, относительное положение образцов в двух группах при t30 было достоверно различным (p < 0.05), но только группа EB достоверно отличалась от образцов при t0 (p < 0.05). Корреляционный график (Рисунок 5B) свидетельствует о том, что наиболее значимой молекулой в структуре, лежащей в основе данных, был сукцинат. Сукцинат был единственной количественно определенной молекулой, у которой корреляция между концентрацией и важностью для PC1 превышала 0.5. Однофакторные сравнения между образцами, взятыми из толстой и подвздошной кишок при t30, предоставили дополнительные возможности для выявления особенностей двух диет. Образцы толстой кишки из группы EB отличались от образцов из группы WB концентрацией семи молекул (дополнительная таблица S6). Модель RPCA, основанная на этих молекулах, позволила четко различать (p < 0.01) образцы из двух групп (Рисунок 5C), при этом валерат и триптофан были более концентрированы в группе WB, а формиат, урацил и ксантин - в группе EB (Рисунок 5D). Параллельный анализ содержимого подвздошной кишки показал, что концентрация восьми молекул в двух группах различалась (Рисунок 5E,F; Дополнительная таблица S7), при этом лактат, 3-метилгистидин и молекула X-5.779 были более концентрированы в группе WB, а 2-гидрокси-3-метилвалерат, трегалоза, 2- оксоглутарат, 2-оксобутират и 5-аминолевулинат более концентрированы в группе EB.
Метаболомика летучих фракций
Чтобы описать влияние двух экспериментальных сортов хлеба (WB и EB) на содержание летучих метаболитов в фекалиях, около 90 летучих молекул были проанализированы методом твердофазной микроэкстракции, газовой хроматографии и масс-спектрометрии (SPME-GC-MS) в образцах фекалий до и после 30-дневной диеты. На рисунке 6 показано распределение летучих соединений до и после вмешательства. Эти две диеты оказали противоположное влияние на разнообразие метаболитов: в частности, диета WB привела к снижению распределения метаболитов, в то время как диета EB привела к увеличению разнообразия метаболитов у экспериментальных животных. Основные метаболитные изменения летучих молекул, полученные в результате выбора признака случайного леса (RF) и последующего корреляционного теста Пирсона, представлены на рисунке 7. В частности, диета WB определяла снижение содержания 2-октена и 2-бутанона (p = 0.005 для обеих молекул; Wilcoxon test) и увеличение концентрации 1-нонанола (p = 0.005). С другой стороны, диета с EB привела к снижению содержания ацетона и 1-октен3-ола (p = 0.005 и 0.013, соответственно), и увеличению содержания пропилового эфира пентановой кислоты (p = 0.065) (Рисунок 7). Частота ошибок из-за отсутствия данных (OOB) составила 0% для WB и 16,67% для EB. Интересно, что среди 10 наиболее важных молекул, которые отвечают за различение между t0 и t30, диета WBхарактеризовалась наличием четырех кетонов, в то время как для диеты EB кетонов было шесть (дополнительный рисунок S3). Среди 10 молекул, идентифицированных с помощью RF, различающих WB и EB в конце исследования (т.е. t30), две молекулы, 1Н-индол-3-метил и 2-тетрадеканол, показали корреляцию, превышающую 0,7 (Корреляция Пирсона показана на дополнительном рисунке S4). Концентрация обеих молекул была выше после диеты WB по сравнению с диетой EB, но только для 1Н-индол-3-метила разница была значительной, при значении p = 0.005 (Рисунок 8).
Обсуждение
Целью настоящего исследования было изучить на модели свиней влияние пшеничного хлеба (WB) и хлеба из однозернянки (EB) на микробиоту и структуру метаболома, как в кале, так и в ЖКТ, а также на гликемические и инсулинемические реакции. Согласно нашим результатам, оба вида хлеба вызывали в целом сходную гликемическую реакцию, в то время как инсулинемическая реакция с точки зрения инсулинового индекса и нагрузки была выше после потребления EB. Поскольку содержание углеводов в хлебе было очень схожим, это различие можно было бы объяснить более высоким содержанием белка в EB по сравнению с WB [34]. Однако, как ранее подчеркивали Abdel-Aal и Hucl [35], наряду с более высоким содержанием белка, продукты из однозернянки содержат относительно мало лизина и много глутаминовой кислоты по сравнению с другими продуктами из пшеницы. Более того, соотношение глиадина и глютенина в белках однозернянки составляло 2:1 по сравнению с 1:1 в пшенице, что указывает на некоторые различия в аминокислотном составе этих сортов пшеницы. Другие различия, которые были обнаружены в EB, включали значительно более высокое содержание липидов, токолов, лютеина и MUFA и более низкое содержание SFAs и PUFAs [36]. Несмотря на это, анализ инсулиновых кривых показал более низкий пик EB, что свидетельствует о более низком и постоянном высвобождении инсулина с течением времени, сравнимом с диетами, которые способствуют длительному чувству сытости [37]. Эта гипотеза, по-видимому, подтверждается снижением гликемического пика, вызванного потреблением EB (Рисунок 1D), что должно вызывать менее интенсивную гипогликемическую реакцию. Интересно, что прием EB привел к двухфазной реакции на гликемию и инсулинемию. Двухфазная форма была коррелирована с более низким риском развития сахарного диабета 2 типа как у подростков, так и у взрослых [38], а в последнее время и с более низкой совокупной заболеваемостью сахарным диабетом 1 типа [39]. В исследовании, проведенном на модели свиньи пероральным тестом на толерантность к глюкозе (OGTT), было показано, что распространенность двухфазных гликемических волн, как и у людей, коррелирует с женским полом [40]. В нашем исследовании все животные, участвовавшие в Tr1, были самками. Мы не исследовали механизм действия, который привел к двухфазной форме, но поскольку исследование было разработано для уменьшения индивидуального эффекта (рисунок в виде латинского квадрата), мы можем обоснованно предположить, что двухфазная форма была вызвана EB. Для изучения этого феномена потребуются дальнейшие исследования. Что касается физиологии животных, то в течение 30-дневного периода интервального питания (Tr2) не наблюдалось существенных изменений показателей крови, за исключением физиологического увеличения гематокрита, гемоглобина, эритроцитов и ретикулоцитов, что соответствует нормальному развитию животных [28]. Прирост массы тела за первые две недели исследования был линейным для обоих рационов и был сопоставим со стандартами для свиней [41]. И наоборот, в обеих группах он замедлился с t14 до t30. Учитывая, что для экспериментальных нужд животные получали одинаковое количество корма (1 кг корма для свиней) в течение всех 30 дней исследования, мы утверждали, что, начиная с периода т14, этого количества было уже недостаточно для поддержания высоких темпов роста, характерных для товарных гибридных свиней. Более того, с t0 по t14 у животных в обеих группах развилась умеренная самоограничивающаяся диарея, скорее всего, вызванная сменой рациона. Действительно, сообщалось, что внезапные изменения в рационе питания могут рассматриваться как фактор пищевого стресса, который может нарушить гомеостаз животных [42], и что пробиотики и противовоспалительные диеты могут помочь уменьшить последствия этих изменений [43]. Интересно, что в группе EB водянистый кал больше не наблюдался со второй недели исследования, что свидетельствует о лучшем и более быстром восстановлении после диареи, связанной с диетическим стрессом, по сравнению с группой WB. Эти результаты согласуются с нашим предыдущим исследованием на культурах кишечных клеток, в котором был выявлен противовоспалительный и защитный эффект при EB, но гематологические показатели воспаления не изменились [4]. Вероятно, мы не смогли обнаружить противовоспалительного эффекта, наблюдая за показателями крови, потому что стресс, вызванный сменой диеты, был очень умеренным и повлиял только на состояние кала. Необходимо провести специальное исследование для изучения противовоспалительного потенциала продуктов на основе однозернянки на модели свиней.
При сравнении с группой, получавшей рацион WB, было обнаружено, что микробиота фекалий свиней, получавших EB, была обогащена несколькими компонентами, которые считались полезными для здоровья, включая хорошо известных продуцентов SCFA: Blautia (ацетогенные), Faecalibacterium (бутирогенные), и Oscillospira. Интересно, что недавние данные указывают на то, что Oscillospira является ключевым представителем кишечной микробиоты и отрицательно связана с воспалительными заболеваниями и индексом массы тела, вероятно, из-за выработки бутирата из пищевых гликопротеинов или гликопротеинов хозяина [44]. В частности, было обнаружено, что относительная численность Oscillospira увеличивается при переходе на рацион, богатый белками и жирами [45], что указывает на интересную связь с белковым и липидным профилем EB, которая заслуживает дальнейшего изучения. С другой стороны, многочисленные исследования подтверждают противовоспалительные свойства Faecalibacterium, широко признанных в качестве биомаркера здоровья кишечника [46], а также Blautia, которые были связаны с уменьшением воспаления и улучшением исходов в нескольких клинических условиях, включая кишечные заболевания, резистентность к инсулину и ожирение, а также иммунологические нарушения [47]. Также стоит отметить, что только диета с EB привела к увеличению относительной распространенности Paludibacter, продуцирующего пропионат и ацетат вида Bacteroidetes [48], и Anaerovibrio, комменсального микроорганизма свиней, известного своей активностью липазы [49,50], что может подтверждать прямую роль этого рода бактерий в развитии снижение высокого содержания липидов в EB по сравнению с WB. Микробиота фекалий свиней, получавших EB, также показала снижение относительной численности семейства Bacteroidales S24-7, которое включает представителей, продуцирующих липополисахариды (LPS), роль которых в физиологии хозяина до сих пор неясна, но доказано, что они способны к оппортунистической инфекции при соответствующих условиях [51,52]. Не в последнюю очередь, на уровне подвздошной кишки фракция микробиоты, ассоциированная со слизистой оболочкой, после 30-дневного приема EB продемонстрировала увеличение биоразнообразия, что является общей мерой, предложенной в качестве показателя здоровья, связанного с эубиозом [53]. Таким образом, возникает соблазн предположить, что потребление EB может модулировать микробный профиль кишечника в сторону изменения конфигурации, которая менее подвержена воспалению, в частности, обогащена микробами, потенциально способствующими поддержанию гомеостаза организма хозяина.
Появляется все больше свидетельств того, что молекулы, присутствующие в просвете кишечника в результате поступления с пищей или продуцируемые кишечной микробиотой, могут играть активную роль в гомеостазе глюкозы. В частности, доступная в кишечнике глюкоза может быть ключевым фактором центрального повышения чувствительности всего организма к инсулину [30]. SCFAs, в частности пропионат, ацетат и бутират, могут действовать как регуляторы гомеостаза глюкозы как при попадании в печень [31] так и при образовании в просвете кишечника [32]. Кроме того, сукцинат и лейцин, продуцируемые микробиотой, были указаны в качестве потенциальных улучшителей гомеостаза глюкозы, в частности, сукцинат способен индуцировать глюконеогенез в кишечнике, выступая в качестве субстрата [29,33]. Чтобы понять, какой из этих известных механизмов, возможно, был активен в настоящем исследовании, было проведено исследование с помощью 1H-NMR вышеупомянутых молекул, за которым последовало нецелевое наблюдение за подвздошной кишкой, ободочной кишкой и фекальным метаболомом. Этот двухэтапный подход выявил ключевую роль сукцината как фекальной молекулы, которая в основном повлияла на структуру образцов. Несмотря на то, что концентрация сукцината в кале животных была одинаковой при t0, было обнаружено, что его содержание значительно снижалось при t30 после диеты с EB, что позволяет предположить, что эта молекула усваивалась более эффективно, что потенциально положительно влияло на гомеостаз глюкозы. Мы даже не можем исключить более высокую скорость утилизации сукцината кишечными бактериями [54], о чем свидетельствует большая относительная численность представителей семейства Bacteroidetes, наблюдаемая только после диеты с EB, которые, как известно, используют сукцинатный путь в качестве основного пути образования пропионата [55]. Более того, модель RPCA показала, что диета с EB смогла существенно повлиять на метаболизм кала животных, в то время как диета с WB - нет. Подтверждая эти результаты, метаболомы содержимого подвздошной и толстой кишки показали значительные различия в зависимости от рациона. Среди молекул, в наибольшей степени влияющих на это различие, следует выделить большое значение триптофана, более концентрированного в группе WB, который, как известно, играет ключевую роль в моторной функции тонкой кишки, а также в ряде каскадных процессов, которые опосредуются индоламиндиоксигеназой (IDO).
Анализ методом GC-MS показал совершенно разные метаболические показатели для двух рационов, с более высоким разнообразием метаболитов в образцах фекалий свиней, которых кормили EB по сравнению с WB. Это согласуется с предыдущими данными о древних злаках, касающимися их влияния на микробиоту кишечника и их метаболиты [23]. В частности, по сравнению с WB, летучий метаболом свиней, получавших EB, показал снижение содержания некоторых спиртов, включая 2-тетрадеканол и 1-октен-3-ол, а также 1Н-индол-3-метил (т.е. скатол), которые, как сообщалось, вызывают повреждение тканей. Действительно, индол представляет собой бактериальный метаболит, который образуется из триптофана, содержащегося в рационе WB в высокой концентрации, как сообщалось выше. Более высокие концентрации такого метаболита могут представлять собой противодействие вредному воздействию LPS в печени при диете с высоким содержанием жиров, о чем недавно сообщалось в литературе [56]. Снижение содержания спиртов может свидетельствовать о снижении производства и, следовательно, о снижении риска эндотоксемии, как обсуждалось в других источниках [57,58]. С другой стороны, было обнаружено, что уровни скатола снижаются в задних кишках и фекалиях свиней, которых кормили с повышенным содержанием сбраживаемых углеводов, что приводит к увеличению количества SCFA и снижению pH, скорее всего, в результате уменьшения присутствия бактерий, продуцирующих скатол, и/или изменения метаболизма триптофана в сторону увеличения производства индола [59]. В этой связи стоит отметить, что Lactobacillus, один из первых бактериальных родов, идентифицированный как продуцент скатола [60], был значительно недопредставлен в микробиоте слизистой оболочки свиней, которых кормили EB, по сравнению с WB. Среди наиболее характерных летучих молекул мы также обнаружили повышенное количество пропилового эфира пентановой кислоты, а также этиловых эфиров бутановой и пропановой кислот в рационе на основе EB. В целом, этерификация с помощью бактериального метаболизма применяется в печени, поджелудочной железе и кишечнике для удаления токсичных концентраций метаболитов, включая SCFA [61], что делает их присутствие очевидным следствием их предшествующего производства.
Выводы
Таким образом, на модели свиней мы продемонстрировали благотворное влияние EB на несколько аспектов физиологии организма хозяина, включая выделение инсулина, консистенцию кала, а также профили микробиоты и метаболизма, как в кале, так и в содержимом кишечника. Согласно нашим выводам, потребление EB может снизить AUC первого инсулинового пика, тем самым продлевая чувство сытости. Более того, оно может модулировать экосистему кишечника, как на уровне состава, так и на уровне метаболизма, в направлении создания конфигурации, специально обогащенной полезными для здоровья бактериями и демонстрирующей отчетливые метаболические признаки, потенциально способствующие поддержанию гомеостаза организма хозяина. Использование модели свиньи позволило, в отличие от клинических испытаний на людях, взять образцы слизистой оболочки и содержимого тонкого кишечника, тем самым расширив знания о сложности взаимодействия пищи, микробиоты и хозяина в ЖКТ. Наблюдаемые положительные эффекты могут быть обусловлены синергетическим взаимодействием многих факторов, включая, в частности, процесс ферментации, пищевую матрицу и компоненты муки, которые оказывают воздействие на кишечник. Оценить полезные свойства настоящих продуктов гораздо сложнее, чем при использовании очищенных пищевых добавок, поскольку редко можно установить прямую причинно-следственную связь между одним компонентом. Именно продукты, а не отдельные компоненты составляют рацион питания, и изучение их сложности может лучше отразить их общую роль для здоровья. Хотя необходимы дальнейшие исследования и клинические испытания, представленные здесь результаты представляют собой первый вклад в раскрытие противовоспалительного потенциала продуктов на основе однозернянки.
Дополнительные Материалы: со следующими материалами можно ознакомиться онлайн по адресу http://www.mdpi.com/2072-6643/11/1/16/s1. Дополнительная таблица S1. Стандартный состав рациона свиней (Big 30–80 by CESAC s.c.a); Дополнительная таблица S2. Уровни глюкозы в крови после приема пищи и инсулина в плазме крови после употребления хлеба из однозернянки (EB) или пшеничного хлеба (WB) на модели свиней (n = 4). EB, 50 г хлеба из однозернянки; WB, 50 г пшеничного хлеба; C, 50 г глюкозы в качестве эталонного продукта. Значения выражены как среднее значение ± SD. Разные строчные буквы указывают на существенные различия между рационами (p < 0.05; Kruskal-Wallis test на нормализованные значения); Дополнительная таблица S3. Общий анализ крови (CBC) и масса тела свиней, включенных в эксперимент Tr 2 выраженные как среднее значение ± стандартная ошибка среднего значения (SEM) для двух рационов (WB, пшеничный хлеб, n = 6; EB, хлеб из однозернянки, n = 6). RBCs (эритроциты): красные кровяные тельца; WBCs (лейкоциты): белые кровяные тельца. Разные строчные буквы указывают на существенные различия в пределах каждой группы между t0 и t30 (p < 0.05, Wilcoxon test); Дополнительная таблица S4. Распределение суточных показателей кала по частоте в двух диетических группах (WB, пшеничный хлеб, n = 6; EB, хлеб из однозернянки, n = 6) в течение четырех недель эксперимента Tr 2. 1 = водянистый кал; 0.5 = пушистый кал; 0 = нормальный кал; Дополнительная таблица S5. Концентрация (мМ), выраженная как среднее ± стандартное отклонение молекул, колебания t0–t30 которых в кале статистически различались в группах (WB) и (EB) (p < 0.1). Значения p были рассчитаны с помощью Wilcoxon test; Дополнительная таблица S6. Концентрация (мМ), выраженная как среднее ± стандартное отклонение молекул (p < 0.1) различающихся между группами (WB) и (EB) в содержимом толстой кишки. Значения p были рассчитаны с помощью Wilcoxon test; Дополнительная таблица S7. Концентрация (мМ), выраженная как среднее ± стандартное отклонение молекул (p < 0.1), различающихся между группами (WB) и (EB) в содержимом подвздошной кишки. Значения p были рассчитаны с помощью Wilcoxon test; Дополнительный рисунок S1. Увеличение массы тела после употребления хлеба из однозернянки (EB, n = 6) и пшеничного хлеба (WB, n = 6). Для обоих рационов (WB - черный и EB - красный) масса тела увеличивалась до 15-го дня (t15) при значении p < 0.01 (ANOVA для повторного измерения) а затем достигла фазы плато; Дополнительный рисунок S2. Соотношение Firmicutes и Bacteroidetes в микробиоте кала свиней, которых кормили хлебом из однозернянки и пшеничным хлебом. Графики в рамках, показывающие распределение соотношения Firmicutes и Bacteroidetes в микробиоте кала свиней до (t0) и после 15 (t15) и 30 (t30) дней употребления (WB) и (EB). Значительное снижение этого соотношения было обнаружено в группе, получавшей диету с EB, как при t15, так и при t30 (p ≤ 0.04, Wilcoxon test); Дополнительный рисунок S3. (A) Случайная классификация по методу RF по 10 наиболее важным молекулам, ответственным за различие между t0 и t30 в диетах с WB и EB. (B) Корреляционный тест Пирсона между идентифицированными молекулами для диеты с WB и EB; Дополнительный рисунок S4. (A) Случайная классификация по методу RF 10 наиболее важных молекул, ответственных за различие между диетой с WB и в конце эксперимента, t30. (B) Корреляционный тест Пирсона между идентифицированными молекулами.
Вклад Авторов: F.B. и D.V. провели эксперименты на модели свиньи и анализ глюкозы и инсулина, A.G., D.L.T.S. и L.L. провели метаболомный анализ, S.T. и P.B. провели характеристику микробиоты. M.F., A.B. и M.L.B. разработали исследование. F.B., S.T., A.G., L.L. и D.L.T.S подготовили рукопись, которая была доработана и одобрена всеми соавторами.
Финансирование: Эта работа была профинансирована European Union’s Seventh Framework Program управляемой REA-Research Executive Agency http://ec.europa.eu/research/rea (FP7/2007-2013) в рамках грантового соглашения n.606476.
Конфликт интересов: Авторы заявляют об отсутствии конфликта интересов.
Источники
1. Dinu, M.; Whittaker, A.; Pagliai, G.; Benedettelli, S.; Sofi, F. Ancient wheat species and human health: Biochemical and clinical implications. J. Nutr. Biochem. 2018, 52, 1–9. [CrossRef] [PubMed]
2. Hidalgo, A.; Brandolini, A. Nutritional properties of einkorn wheat (Triticum monococcum L.). J. Sci. Food Agric.
2014, 94, 601–612. [CrossRef] [PubMed]
3. Molberg, Ø.; Uhlen, A.K.; Jensen, T.; Flæte, N.S.; Fleckenstein, B.; Arentz-Hansen, H.; Raki, M.; Lundin, K.E.A.; Sollid, L.M. Mapping of gluten T-cell epitopes in the bread wheat ancestors: Implications for celiac disease. Gastroenterology 2005, 128, 393–401. [CrossRef] [PubMed]
4. Antognoni, F.; Mandrioli, R.; Bordoni, A.; Di Nunzio, M.; Viadel, B.; Gallego, E.; Villalba, M.P.; Tomás-Cobos, L.; Taneyo Saa, D.L.; Gianotti, A. Integrated evaluation of the potential health benefits of einkorn-based breads. Nutrients 2017, 9, 1232. [CrossRef] [PubMed]
5. Turroni, S.; Brigidi, P.; Cavalli, A.; Candela, M. Microbiota-host transgenomic metabolism, bioactive molecules from the inside. J. Med. Chem. 2018, 61, 47–61. [CrossRef] [PubMed]
6. Bordoni, A.; Danesi, F.; Di Nunzio, M.; Taccari, A.; Valli, V. Ancient wheat and health: A legend or the reality? A review on KAMUT khorasan wheat. Int. J. Food Sci. Nutr. 2017, 68, 278–286. [CrossRef] [PubMed]
7. Thorup, A.C.; Gregersen, S.; Jeppesen, P.B. Ancient wheat diet delays diabetes development in a type 2 diabetes animal model. Rev. Diabet. Stud. 2014, 11, 245. [CrossRef]
8. Bo, S.; Seletto, M.; Choc, A.; Ponzo, V.; Lezo, A.; Demagistris, A.; Evangelista, A.; Ciccone, G.; Bertolino, M.; Cassader, M.; et al. The acute impact of the intake of four types of bread on satiety and blood concentrations of glucose, insulin, free fatty acids, triglyceride and acylated ghrelin. A randomized controlled cross-over trial. Food Res. Int. 2017, 92, 40–47. [CrossRef] [PubMed]
9. Graham, H.; Åman, P. The pig as a model in dietary fibre digestion studies. Scand. J. Gastroenterol. 1987, 22,
55–61. [CrossRef]
10. Kararli, T.T. Comparison of the gastrointestinal anatomy, physiology, and biochemistry of humans and commonly used laboratory animals. Biopharm. Drug Dispos. 1995, 16, 351–380. [CrossRef]
11. Leulier, F.; MacNeil, L.T.; Lee, W.J.; Rawls, J.F.; Cani, P.D.; Schwarzer, M.; Zhao, L.; Simpson, S.J. Integrative physiology: At the crossroads of nutrition, microbiota, animal physiology, and human health. Cell Metab. 2017, 25, 522–534. [CrossRef] [PubMed]
12. Bryszewska, M.A.; Laghi, L.; Zannoni, A.; Gianotti, A.; Barone, F.; Taneyo Saa, D.L.; Bacci, M.L.; Ventrella, D.; Forni, M. Bioavailability of microencapsulated iron from fortified bread assessed using piglet model. Nutrients 2017, 9, 272. [CrossRef] [PubMed]
13. Venn, B.J.; Green, T.J. Glycemic index and glycemic load: Measurement issues and their effect on diet–disease relationships. Eur. J. Clin. Nutr. 2007, 61, S122. [CrossRef] [PubMed]
14. De La Fuente-Arrillaga, C.; Martinez-Gonzalez, M.A.; Zazpe, I.; Vazquez-Ruiz, Z.; Benito-Corchon, S.; Bes-Rastrollo, M. Glycemic load, glycemic index, bread and incidence of overweight/obesity in a Mediterranean cohort: The SUN project. BMC Public Health 2014, 14, 1091. [CrossRef] [PubMed]
15. Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples.
Biotechniques 2004, 36, 808–812. [CrossRef] [PubMed]
16. Biagi, E.; Franceschi, C.; Rampelli, S.; Severgnini, M.; Ostan, R.; Turroni, S.; Consolandi, C.; Quercia, S.; Scurti, M.; Monti, D.; et al. Gut microbiota and extreme longevity. Curr. Biol. 2016, 26, 1480–1485. [CrossRef] [PubMed]
17. Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [CrossRef] [PubMed]
18. Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pẽa, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [CrossRef]
19. Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461.
[CrossRef]
20. Ventrella, D.; Laghi, L.; Barone, F.; Elmi, A.; Romagnoli, N.; Bacci, M.L. Age-related 1H NMR characterization of Cerebrospinal fluid in newborn and young healthy piglets. PLoS ONE 2016, 11, e0157623. [CrossRef]
21. Barbara, G.; Scaioli, E.; Barbaro, M.R.; Biagi, E.; Laghi, L.; Cremon, C.; Marasco, G.; Colecchia, A.; Picone, G.; Salfi, N.; et al. Gut microbiota, metabolome and immune signatures in patients with uncomplicated diverticular disease. Gut 2017. [CrossRef] [PubMed]
22. Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in 1H NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [CrossRef] [PubMed]
23. Taneyo Saa, D.; Turroni, S.; Serrazanetti, D.I.; Rampelli, S.; Maccaferri, S.; Candela, M.; Severgnini, M.; Simonetti, E.; Brigidi, P.; Gianotti, A. Impact of Kamut®Khorasan on gut microbiota and metabolome in healthy volunteers. Food Res. Int. 2014, 63, 227–232. [CrossRef]
24. Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [CrossRef] [PubMed]
25. Enot, D.P.; Lin, W.; Beckmann, M.; Parker, D.; Overy, D.P.; Draper, J. Preprocessing, classification modeling and feature selection using flow injection electrospray mass spectrometry metabolite fingerprint data. Nat. Protoc. 2008, 3, 446–470. [CrossRef] [PubMed]
26. Conover, W.J. Practical Nonparametric Statistics, 3rd ed.; John Wiley & Sons: London, UK, 1980; ISBN
0471028673r0471084573.
27. Hubert, M.; Rousseeuw, P.J.; Vanden Branden, K. ROBPCA: A new approach to robust principal component analysis. Technometrics 2005, 47, 64–79. [CrossRef]
28. Ventrella, D.; Dondi, F.; Barone, F.; Serafini, F.; Elmi, A.; Giunti, M.; Romagnoli, N.; Forni, M.; Bacci, M.L. The biomedical piglet: Establishing reference intervals for haematology and clinical chemistry parameters of two age groups with and without iron supplementation. BMC Vet. Res. 2017. [CrossRef] [PubMed]
29. Layman, D.K. Symposium: Dairy product components and weight regulation the role of leucine in weight loss diets and glucose homeostasis 1,2. J. Nutr. 2003. [CrossRef] [PubMed]
30. Mithieux, G. A novel function of intestinal gluconeogenesis: Central signaling in glucose and energy homeostasis. Nutrition 2009, 25, 881–884. [CrossRef] [PubMed]
31. Ulven, T. Short-chain free fatty acid receptors FFA2/GPR43 and FFA3/GPR41 as new potential therapeutic targets. Front. Endocrinol. 2012, 3, 111. [CrossRef] [PubMed]
32. Psichas, A.; Reimann, F.; Gribble, F.M. Gut chemosensing mechanisms. J. Clin. Investig. 2015, 125, 908–917.
33. De Vadder, F.; Kovatcheva-Datchary, P.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-produced succinate improves glucose homeostasis via intestinal gluconeogenesis. Cell Metab. 2016. [CrossRef] [PubMed]
34. Nuttall, F.Q.; Mooradian, A.D.; Gannon, M.C.; Billington, C.J.; Krezowski, P. Effect of protein ingestion on the glucose and insulin response to a standardized oral glucose load. Diabetes Care 1984. [CrossRef]
35. Abdel-Aal, E.S.M.; Hucl, P. Amino acid composition and in vitro protein digestibility of selected ancient wheats and their end products. J. Food Compos. Anal. 2002, 15, 737–747. [CrossRef]
36. Hidalgo, A.; Brandolini, A.; Ratti, S. Influence of genetic and environmental factors on selected nutritional traits of Triticum monococcu. J. Agric. Food Chem. 2009, 57, 6342–6348. [CrossRef] [PubMed]
37. Rosén, L.A.H.; Östman, E.M.; Shewry, P.R.; Ward, J.L.; Andersson, A.A.M.; Piironen, V.; Lampi, A.M.; Rakszegi, M.; Bedö, Z.; Björck, I.M.E. Postprandial glycemia, insulinemia, and satiety responses in healthy subjects after whole grain rye bread made from different rye varieties. 1. J. Agric. Food Chem. 2011. [CrossRef]
38. Bervoets, L.; Mewis, A.; Massa, G. The shape of the plasma glucose curve during an oral glucose tolerance test as an indicator of beta cell function and insulin sensitivity in end-pubertal obese girls. Horm. Metab. Res. 2015. [CrossRef]
39. Ismail, H.M.; Xu, P.; Libman, I.M.; Becker, D.J.; Marks, J.B.; Skyler, J.S.; Palmer, J.P.; Sosenko, J.M. The shape of the glucose concentration curve during an oral glucose tolerance test predicts risk for type 1 diabetes. Diabetologia 2018. [CrossRef]
40. Manell, E.; Hedenqvist, P.; Svensson, A.; Jensen-Waern, M. Establishment of a refined oral glucose tolerance test in pigs, and assessment of insulin, glucagon and glucagon-like peptide-1 responses. PLoS ONE 2016. [CrossRef]
41. Carr, J. Garth Pig Stockmanship Standards; 5M Enterprises Limited: Sheffield, UK, 1998.
42. Suiryanrayna, M.V.A.N.; Ramana, J.V. A review of the effects of dietary organic acids fed to swine. J. Anim. Sci. Biotechnol. 2015, 6, 45. [CrossRef]
43. Respondek, F.; Goachet, A.G.; Julliand, V. Effects of dietary short-chain fructooligosaccharides on the intestinal microflora of horses subjected to a sudden change in diet. J. Anim. Sci. 2008, 86, 316–323. [CrossRef] [PubMed]
44. Konikoff, T.; Gophna, U. Oscillospira: A central, enigmatic component of the human gut microbiota.
Trends Microbiol. 2016, 24, 523–524. [CrossRef] [PubMed]
45. David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [CrossRef] [PubMed]
46. Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841. [CrossRef]
47. Jenq, R.R.; Taur, Y.; Devlin, S.M.; Ponce, D.M.; Goldberg, J.D.; Ahr, K.F.; Littmann, E.R.; Ling, L.; Gobourne, A.C.; Miller, L.C.; et al. Intestinal blautia is associated with reduced death from graft-versus-host disease. Biol. Blood Marrow Transplant. 2015. [CrossRef]
48. Qiu, Y.L.; Kuang, X.Z.; Shi, X.S.; Yuan, X.Z.; Guo, R.B. Paludibacter jiangxiensis sp. nov., a strictly anaerobic, propionate-producing bacterium isolated from rice paddy field. Arch. Microbiol. 2014. [CrossRef]
49. He, M.; Fang, S.; Huang, X.; Zhao, Y.; Ke, S.; Yang, H.; Li, Z.; Gao, J.; Chen, C.; Huang, L. Evaluating the contribution of gut microbiota to the variation of porcine fatness with the cecum and fecal samples. Front. Microbiol. 2016. [CrossRef]
50. Holman, D.B.; Brunelle, B.W.; Trachsel, J.; Allen, H.K. Meta-analysis to define a core microbiota in the swine gut. mSystems 2017. [CrossRef]
51. Ormerod, K.L.; Wood, D.L.A.; Lachner, N.; Gellatly, S.L.; Daly, J.N.; Parsons, J.D.; Dal’Molin, C.G.O.; Palfreyman, R.W.; Nielsen, L.K.; Cooper, M.A.; et al. Genomic characterization of the uncultured Bacteroidales family S24-7 inhabiting the guts of homeothermic animals. Microbiome 2016. [CrossRef]
52. Kang, C.; Wang, B.; Kaliannan, K.; Wang, X.; Lang, H.; Hui, S.; Huang, L.; Zhang, Y.; Zhou, M.; Chen, M.; et al. Gut microbiota mediates the protective effects of dietary capsaicin against chronic low-grade inflammation and associated obesity induced by high-fat diet. MBio 2017. [CrossRef]
53. Sonnenburg, E.D.; Sonnenburg, J.L. Starving our microbial self: The deleterious consequences of a diet deficient in microbiota-accessible carbohydrates. Cell Metab. 2014, 20, 779–786. [CrossRef] [PubMed]
54. Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol.
2017, 19, 29–41. [CrossRef] [PubMed]
55. Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014. [CrossRef] [PubMed]
56. Beaumont, M.; Neyrinck, A.M.; Olivares, M.; Rodriguez, J.; de Rocca Serra, A.; Roumain, M.; Bindels, L.B.; Cani, P.D.; Evenepoel, P.; Muccioli, G.G.; et al. The gut microbiota metabolite indole alleviates liver inflammation in mice. FASEB J. 2018, fj.201800544. [CrossRef] [PubMed]
57. Cani, P.D. Gut microbiota and pregnancy, a matter of inner life. Br. J. Nutr. 2009, 101, 1579–1580. [CrossRef]
[PubMed]
58. Vernocchi, P.; Del Chierico, F.; Putignani, L. Gut microbiota profiling: Metabolomics based approach to unravel compounds affecting human health. Front. Microbiol. 2016, 7, 1144. [CrossRef] [PubMed]
59. Vhile, S.G.; Kjos, N.P.; Sørum, H.; Øverland, M. Feeding Jerusalem artichoke reduced skatole level and changed intestinal microbiota in the gut of entire male pigs. Animal 2012, 6, 807–814. [CrossRef] [PubMed]
60. Yokoyama, M.T.; Carlson, J.R.; Holdeman, L.V. Isolation and characteristics of a skatole-producing
Lactobacillus sp. from the bovine rumen. Appl. Environ. Microbiol. 1977, 34, 837–842.
61. Vitali, B.; Ndagijimana, M.; Cruciani, F.; Carnevali, P.; Candela, M.; Guerzoni, M.E.; Brigidi, P. Impact of a synbiotic food on the gut microbial ecology and metabolic profiles. BMC Microbiol. 2010, 10, 4. [CrossRef]