Что лежит в ящике с инструментами генного инженера?
Любая современная технология требует использования высокоточных инструментов для ее реализации. И если для тяжелой промышленности нужны мощные лазеры и роботизированные станки (и еще много чего другого), то для биотехнологии ключевую роль играют специализированные молекулярные инструменты, позволяющие аккуратно вносить изменения в генетический материал организма-мишени. К счастью, большую часть работы в этом направлении за нас сделала эволюция, снабдив вирусы разнообразными белками (или кодирующими их генами), которые помогают не только успешно попасть в организм «хозяина», но и встроить свою генетическую информацию в его геном.
Другие инструменты, а именно ферменты, способные разрезать или модифицировать чужеродный генетический материал, появились в клетках бактерий и некоторых эукариот в ответ на вирусную угрозу. Эти инструменты совершенствовались миллиарды лет, а мы знаем о них всего несколько десятилетий. Тем не менее, применение этих инструментов в биотехнологии уже позволило добиться серьезных успехов, и ниже я расскажу об основных особенностях этого молекулярного арсенала. Методы генной инженерии мы уже разбирали подробно в материалах спецпроекта «12 биологических методов в картинках»; тут приведем лишь краткий обзор этих подходов, который поможет вам освежить в памяти основные достижения и современные технологии, лежащие в основе генной инженерии.
Инструменты для редактирования генома
Более подробно эта тема раскрыта в статьях «12 методов в картинках: генная инженерия. Часть I, историческая» и «...Часть II: инструменты и техники», очень рекомендуем к прочтению!
Рекомбиназы —
это ферменты, вызывающие сайт-специфический мутагенез. Они распознают определенные последовательности ДНК и вызывают обмен между участками ДНК — то есть, рекомбинацию. Именно эта технология стала основой для появления первых генномодифицированных животных и в сочетании с использованием эмбриональных стволовых клеток сделала возможной доставку трансгена и появление первых успешных генетических моделей.
Один из первых подобных инструментов — система Cre-LoxP, характерная для бактериофага P1, открытая и приспособленная для использования на животных еще в далекие (по меркам биотеха) 1980-е годы. Она состоит из двух основных компонентов: (1) фермент Cre, который может распознавать (2) специфичный для него участок ДНК LoxP длиной 34 пары нуклеотидов, вызывая в нем разрыв и рекомбинацию. Характер расположения сайтов LoxP может вызывать разнообразные изменения в ДНК: делеции, инсерции, вставки и рекомбинацию фрагментов молекулы ДНК. Механизм внесения изменений в участке ДНК показан на схеме ниже. Как правило, рекомбиназа Cre распознает сайты LoxP, ограничивающие некий фрагмент ДНК, и создает петлю, которую затем можно инвертировать или вырезать в зависимости от ориентации сайтов LoxP (более подробное объяснение в подписи к рисунку).
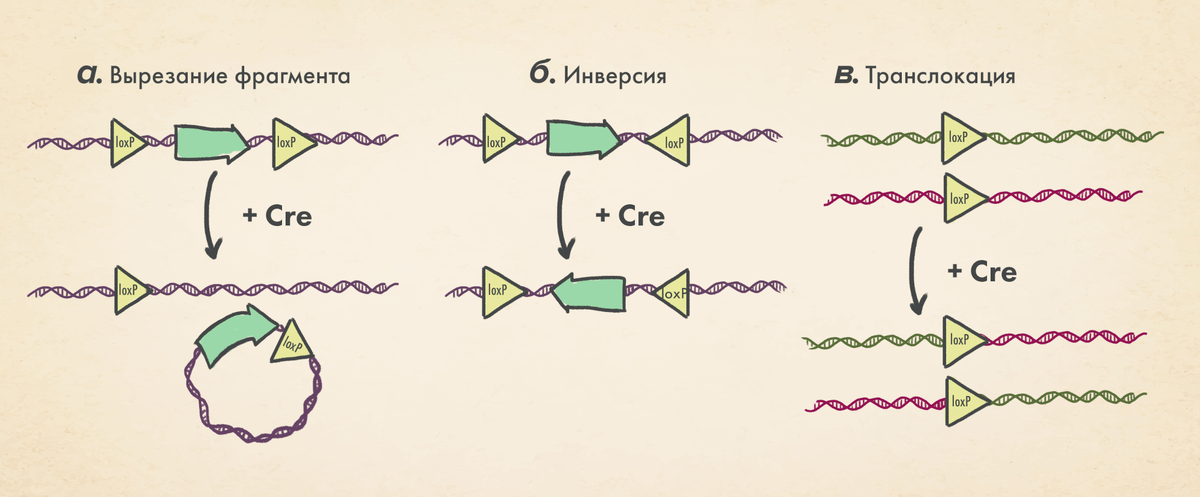
Технология Cre-LoxP позволяет вносить изменения в генетический аппарат клеток заранее заданного типа, сильно выигрывая у генной модификации всех клеток организма. Чтобы успешно применять эту технологию на животных (например, на мышах), необходимо иметь две линии: одна из них должна будет содержать последовательности LoxP, а другая — ген рекомбиназы Cre. Причем экспрессия гена Cre должна управляться определенным тканеспецифичным промотором, чтобы рекомбиназа появлялась только в определенных клетках-«мишенях». Далее, необходимо скрещивать мышей до тех пор, пока оба компонента системы не встретятся в целевой ткани. Тогда произойдет рекомбинация, приводящая к тому событию, на которое рассчитывает исследователь: «выключение» (knock out) или, наоборот, «включение» (knock in) гена.
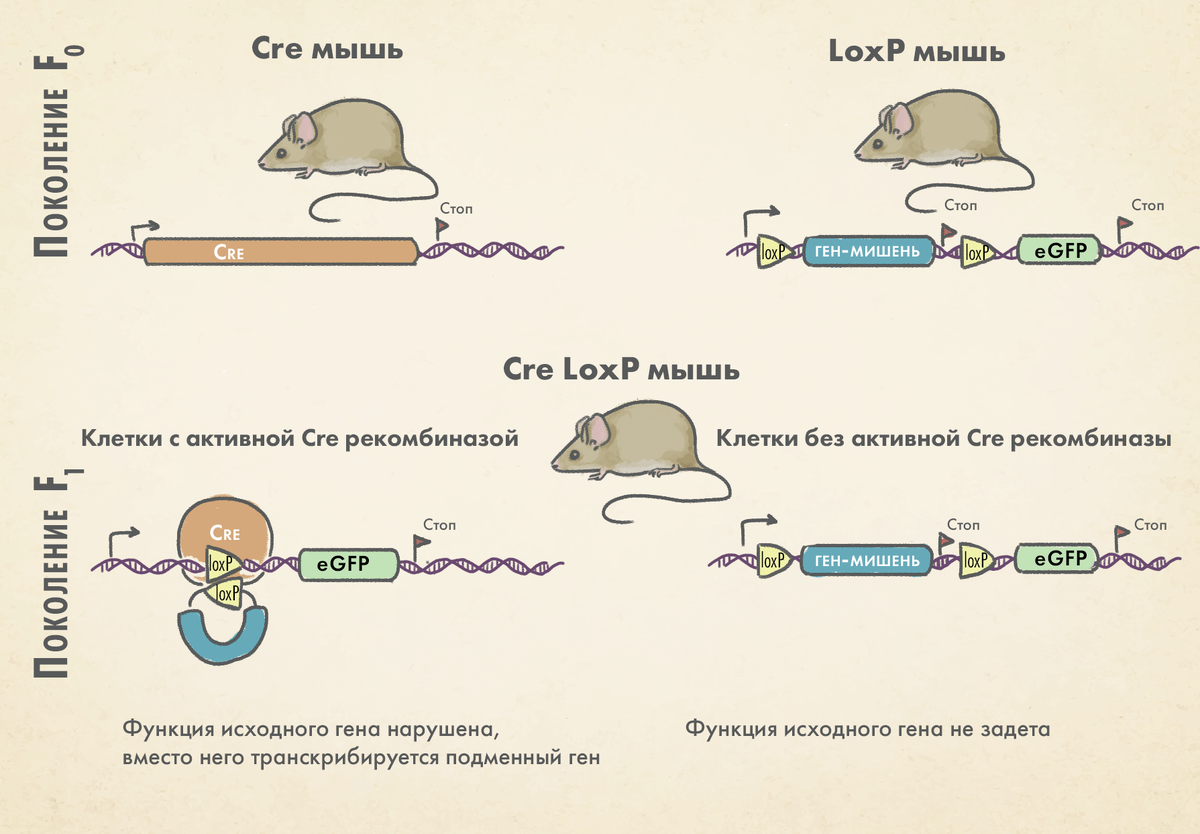
Задачи современной генной инженерии становятся все сложнее, и одного сайта LoxP не хватает для построения необходимых экспериментатору моделей. Поэтому технологии на основе Cre-LoxP продолжали развиваться. Появлялись новые последовательности VloxP и SloxP, распознаваемые ферментом Cre, а также открыты новые рекомбиназы. Ферментов, аналогичных Cre, в природе немало; некоторые источники сообщают о более чем 1300 подобных рекомбиназ. Но к сайтам, которые распознают рекомбиназы, предъявляются высокие требования. Так, например, они не должны вызывать рекомбинацию на случайных участках ДНК клетки хозяина. Также в настоящее время появились новые рекомбиназы Nigri и Panto, расширяющие арсенал инструментов редактирования генома.
Нуклеазы —
это ферменты, которые вносят двухцепочечные разрывы в определенных участках ДНК, вызывая необходимые модификации в нужном гене. В отличие от рекомбиназ, которые работают только в процессе репликации (удвоения) ДНК, нуклеазы действуют на любом этапе клеточного цикла, что значительно расширяет возможности их использования. Открытие настраиваемых нуклеаз, которые можно запрограммировать на индукцию двухцепочечных разрывов ДНК в желаемых локусах генома, значительно повысило эффективность рекомбинации, что привело к еще одной революции в редактировании генов.
Ниже рассмотрим основные типы нуклеаз, используемых в настоящее время для создания моделей трансгенных животных.
ZFN
имеют в своем составе домены, взаимодействующие с молекулой ДНК и напоминающие по форме прищепку, а в качестве «заклепки» выступает ион цинка. Поэтому подобный белковый мотив называют цинковым пальцем, что впоследствии дало название всей нуклеазе: Zinc-Finger Nuclease (ZFN). Это димер, мономер которого имеет в своем составе три мотива с цинковым пальцем и эндонуклеазный домен Fokl, разрезающий ДНК. За счет цинковых мотивов можно «нацелить» эндонуклеазу на нужный участок ДНК и внести разрыв, который затем сшивается с помощью механизмов репарации, описанных выше. Это приводит к изменению аминокислотной последовательности белка, кодируемого целевым геном, и поэтому используется как способ создания точечных мутаций в самых разнообразных организмах от дрозофилы до мыши или крысы.
ZFN также применяли для создания мутации в хемокиновом рецепторе CCR5, который ответственен за проникновение ВИЧ в клетку. Появление в арсенале генных инженеров нуклеазы ZFN позволило значительно расширить возможности редактирования генома и создать первые трансгенные модели крыс, дрозофил, некоторых рыб, чего не удавалось раньше с использованием рекомбиназ. Однако к минусам этой технологии можно отнести сложность в оптимизации ZFN под целевую последовательность и внесение двунитевых разрывов в нецелевые участки ДНК. Сложность, например, может состоять в том, что один цинковый палец может влиять на соседний, что порой сложно учитывать при проектировании специфичных мотивов.
TALEN (Transcription Activator-Like Effector Nuclease) —
эндонуклеаза, похожая на ZFN: повторяющиеся мотивы для связывания ДНК (TALE) и эндонуклеазу, подобную Fokl. Однако каждый мотив TALE связывается с отдельным нуклеотидом, что позволяет более точно «настраивать» эндонуклеазу. Две пары TALEN можно использовать для создания делеций. Преимуществом TALEN перед ZFN можно назвать бóльшую специфичность к целевому участку ДНК благодаря гибкой настройке мотивов TALE и более низкий процент нецелевых эффектов нуклеазы. Минусами этой технологии является большой размер TALEN, из-за которого сложнее найти вектор подходящей вместительности. Кроме того, опасения специалистов вызывает повышенная иммуногенность эндонуклеазы. Появление TALEN продолжило путь нуклеаз для создания трансгенных моделей. Несмотря на то, что рекомбиназы уже давно используются для создания трансгенных мышей и некоторых других животных, такие нуклеазы, как TALEN, делают это быстрее, дешевле и эффективнее.
CRISPR/Cas9 —
система редактирования генома прокариот, которая приобрела огромную популярность в генной инженерии в последнее десятилетие [30]. В основе системы лежит использование РНК-затравки CRISPR (crRNA) и трансактивирующей РНК (tracrRNA), формирующих сайт, распознаваемый нуклеазой Cas9, которая после распознавания может разрезать чужеродные генетические элементы (рис. 5). crRNA можно заменить любой целевой последовательностью, комплементарной определенному участку ДНК, и для этого используют РНК, заменяющую CRISPR и трансактивирующую РНК химерной РНК (sgRNA).
Эта система редактирования широко используется как на клетках животных, так и человека для внесения и исправления генетических мутаций [31]. Система CRISPR/Cas9 удобнее в работе, чем TALEN и ZFN: нужно лишь заменить последовательность sgRNA на комплементарную целевому участку ДНК, в то время как сам фермент Cas9 останется без изменений. Появление системы CRISPR/Cas9 произвело настоящую революцию в генной инженерии. Создание трансгенных моделей животных с технологией CRISPR/Cas9 вышло на новый уровень по ряду причин:
- В отличие от других систем, CRISPR/Cas9 применима в широком диапазоне биологических видов. Это связано с тем, что библиотеки CRISPR можно создавать по всему геному любого биологического вида. Необходимая для работы sgRNA комплементарна целевому участку ДНК, а значит может быть подобрана практически для любого фрагмента ДНК. В отличие от CRISPR/Cas9, системы ZFN и TALEN нельзя оптимизировать для любой последовательности ДНК (это связано с особенностями взаимодействий ДНК–белок), что ограничивает их применение.
- Систему CRISPR/Cas9 легко запрограммировать на множество локусов в одной клетке, что позволяет использовать технологию для скрининга генов-кандидатов для лечения ряда заболеваний. Подобный скрининг использовался на моделях мышей для поиска генов, которые могут играть важную роль в терапии рака.
- Появление технологии CRISPR/Cas9 значительно ускорило создание трансгенных моделей заболеваний человека. Создание точечных мутаций, крупных делеций или даже перенос целых генов человека и создание гуманизированных животных стало рутинной процедурой. Однако использование CRISPR/Cas9 имеет свои побочные эффекты, в том числе появление антител к определенным изоформам Cas9 и другие неспецифические реакции со стороны клетки.
Больше о том, как внедрять генетические конструкции в клетки и как создаются и для чего используются трансгенные животные, читайте на нашем сайте!