
ФАКТ 14
Инстинктивный ответ можно вызвать грубой моделью, совсем непохожей на «нужное» животное-компаньона, лишь бы несла «нужный» знак в «нужной» позиции. Скажем, агонистические демонстрации трёхиглых колюшек вызывают модели, не имеющие ничего общего с рыбкой, лишь бы красные снизу. И наоборот, точная копия рыбки, но без красного низа, вовсе неэффективна (рис.16А) — считали классические этологи.
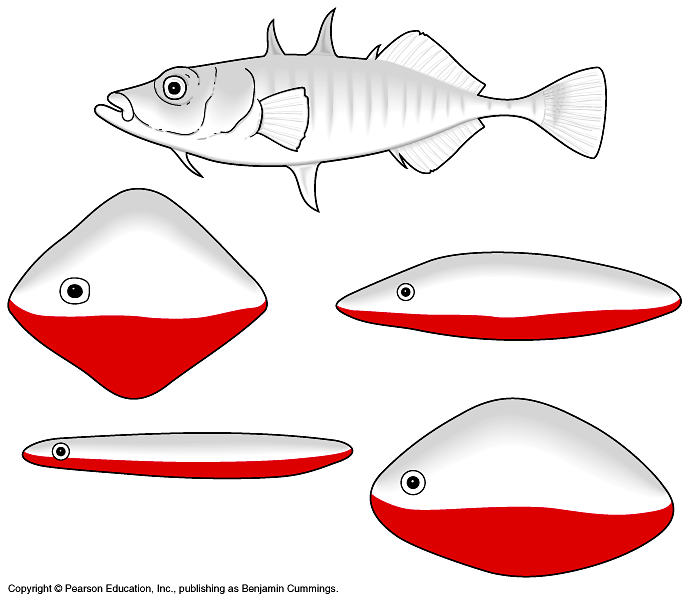
А.
Б.
Однако этот вывод основан лишь на отдельных «парадных примерах». Как ни странно, до середины 1980-х гг. его так и не подкрепили контролируемым экспериментом со статистической обработкой данных, показывающим существование (или нет) значимых различий между красной снизу vs серой моделями по частоте ударов от самца колюшки. Необходимые опыты провёл лишь Nicolas E. Collias в 1990 г.
Р. Sevenster (1951) показал, что агрессивность самца при строительстве гнезда[60] растёт со временем, выходя на плато по его завершении (тогда самцов и тестировали); она максимальна вблизи гнезда и монотонно снижается с удалением от него. Поэтому в опытах 10 самцам А-J подавали восковую модель рыбки естественного размера и формы на проволоке, крепящейся к её спине, на дистанции 7-13 см от гнезда. Другой конец проволоки крепили сбоку баков, где жили самцы. Один вариант модели был красным, другой серым снизу, оба предъявляли горизонтально, т. е. в нейтральной позе: положение «головой внизу» у колюшек — поза угрозы.
При каждом тестировании считали число укусов, полученных моделью за 5 мин наблюдений. За раз предъявляли только одну из альтернатив: после каждого предъявления ждали 10 мин до следующего. Предъявления «красной» и «серой» моделей разным самцам чередуются, как показано на рис.16Б, чтобы каждая из них была первой или второй равное число раз. Большинство предъявлений сделаны утром, если была вторая серия опытов с тем же самцом — вечером, чтобы избежать привыкания к стимулу.
Данные подтвердили вывод о релизерном эффекте «красного низа» модели, самцы колюшек кусали его значимо чаще, чем серый (рис.16Б). Классические выводы, однако, пришлось скорректировать:
а) значительной вариабельностью ответа между самцами, включая его отсутствие у А (возможно, из-за загруженности «родительскими заботами», у него было 5 кладок). Самец Н (одна кладка) интенсивно бросался на красную снизу модель, но почти не кусал её. Прочие самцы только выстроили гнездо, но не обладали кладками;
б) при значимости различий в реакции на обе модели у 4 из 9 рыб разница была невелика;
в) самцы атакуют и серую снизу модель, хоть и значимо реже: возможно, вследствие сочетания растущего возбуждения и длительной депривации, снижающей порог реагирования, т. е. кусание серой модели — просто активность вхолостую.
Ряд авторов до него также сообщали о предпочтительной реакции самцов колюшек на красные снизу модели, сравнительно с серыми снизу или с иной окраской, помимо красного. Эти случаи (включая сообщённые N.E.Colias) рассмотрел G.Baerends (1985), но без статистического анализа. Они оказались противоречивы, возможно из-за неконтролируемости побуждений к бегству, возникающих у самца из-за перемещений модели. Чтобы избежать этого, N.E. Colias (1990) помещал неподвижную модель в центре территории.
Вернёмся к ежам. Во взаимодействии резидента с захватчиком демонстрации и прямая агрессия чередуются, за демонстрацией следует «выплеск» действий открытого нападения[61]. Если взаимодействие включает коммуникацию, агрессия особей в отношении друг друга происходит не напрямую, но опосредована её ритуализованным выражением, демонстрацией. На агрессию атакуемый отвечает агрессию не бегством или контратакой, но ответной демонстрацией: он маневрирует собственным колючим панцирем, в описанной манере сдвигая кожу спины в сторону грозящей опасности. Видим, что именно демонстрация заставляет успешно атакующего противника остановиться и самому начать демонстрировать, то есть перейти от прямого действия к обмену сигналами, выявляющему победителя «по правилам системы». Действительно, если один из зверьков перестаёт демонстрировать и обращается в бегство, нападающий теряет к нему интерес и перестаёт преследовать.
Когда взаимодействие осуществляется некоммуникативным способом, ответ открытой агрессией следует не только на аналогичную агрессию, но и на демонстрации оппонента; такой процесс перерастает в открытую драку, в которой не различить победителя и побеждённого (Карасёва, Готфрид, 1983).
ФАКТ 15
Отсюда естественен вывод о «сигнальности» или «знаковости» релизеров. Хотя их «зовут» знаковыми стимулами, прилагательное здесь несравнимо важней существительного. Стимул исходно заострённая палка, непосредственно (уколом, тычком или толчком) «принуждающая» к определённым ответам.
Так, при ухаживании самка зеленоватого тритона Notophtalmus viredescens демонстрирует брюхо, заполненное яйцами, самец ей — внутренние поверхности задних конечностей с роговыми подушечками, небольшие ямки на скулах и хвост с увеличенной высокой складкой. Реакция самки на это задаёт два сценария поведения самца: если она остановится рядом с ним, демонстрирование быстро закончится и самец перейдёт к следующей фазе ухаживания. Если она удаляется, самец следует за ней, стараясь обхватить передними лапками, и интенсивно демонстрирует. При низкой реактивности самки продолжительность демонстраций примерно в 25 раз выше: в конце концов, движение самки прочь замедляется, самцу удаётся её нагнать и схватить[62].
Увеличивая интенсивность воздействия стимула, самец «поднимает» реактивность самки до уровня, позволяющего ей перейти к следующим этапам ухаживания – сближению с демонстрирующими самцами, чтобы выбрать одного и взаимодействовать дальше именно с ним вплоть до передачи сперматофора. Интенсивность воздействия при таком «обращении внимания» самки на самцов пока что важней точности воспроизведения форм демонстраций. Не случайно она ведёт к характерным «сбоям» — привлечению незрелых самок, самок близкого вида и т.п. «посторонних объектов», негодных для ухаживания. Когда же под действием соответствующей стимуляции самка уже мотивирована к сближению с самцами и к оценке их демонстраций «с точки зрения» последующей копуляции, точность воспроизведения их специфических форм делается решающей, а интенсивность стимуляции не столь важна. См. далее про ухаживание девятииглых колюшек рода Pungitius.
Так или иначе, релизер действует своей специфической формой: даже когда «принуждающее» воздействие присутствует, оно реализуется лишь при условии, что: а) предъявление демонстрации-релизера достаточно стереотипно, т. е. образующие её телодвижения «чертят» образ, воспринимаемый реципиентом, так, что он инвариантен к изменениям контекста; б) этот образ устойчиво воспринят реципиентом вопреки разнообразным «помехам», внешним и внутренним.
Так, исследовали индикаторы эмоционального возбуждения в структуре оборонительных криков большой песчанки Rhombomys opimus, и их действие на агрессора. Конфликт провоцировали, ссаживая зверьков на нейтральной территории. После короткого периода борьбы, в разной степени ритуализованной, один (победитель) преследовал второго, проявлявшего подчинённое поведение и издававшего оборонительные крики. Сокращение дистанции между ними меняло эмоциональное состояние обоих, что сразу отображалось в пропорциональных изменениях вокализации: падала длительность оборонительных звуков, их энергия смещалась в область высоких частот. Для тональных звуков выросли значения основной частоты и глубины частотной модуляции.
Данные изменения в криках испуганной, обороняющейся песчанки сразу «отталкивали» наступающую, резко увеличивая дистанцию между ними. Важно, что крики больших песчанок, обладающие более специализированной акустической формой, эффективней ингибировали агрессию по сравнению с менее определёнными криками светлой Gerbillus perpallidus[63] (Володин и др., 1998; Доронина, Володин, 2007).
Третий пример двойственного действия релизеров — как сигналов и специфических стимулов, где второе реализуемо лишь при условии первого, но не наоборот, даёт булькающая вокализация черноголовых гаичек (рис.17А). С одной стороны, бульканье — внешнее выражение агрессивности, т. е. способности птицы силой очистить кормушку, заставить товарок отступить и ждать своей очереди. С другой, его акустическая форма — предиктор большей или меньшей способности изгонять других особей, в «сквозной» оценке которой «заинтересованы» все совместно кочующие члены зимних стай, а также бродячие птицы (Baker et al., 1991).
Анализ показывает, что бульканье связано со стремлением птиц доминировать в конкурентной ситуации, скажем, согнать другую или брать корм, не обращая ни на кого внимания. В конфликтах между самцами гаичек всегда побеждает булькающий (рис.17Б). При ссаживании незнакомых птиц на нейтральной территории доминирование устанавливается посредством борьбы. В её начале гаички обмениваются булькающими сигналами, чередуя их с открытой агрессией по отношению к оппоненту (Baker et al., 1991). «Бульканье» здесь — достоверный маркер агрессивности, почему выступает сигналом состояния, предиктором того «количества агрессии», которое животное может «обрушить» для вытеснения или подчинения данной особи.
В какой-то момент одна гаичка прекращает сопротивление, а вторая может её изгонять просто бульканьем, без открытой агрессии: здесь появляется доминирование. В устойчивых зимних стаях синиц бульканье — знак присутствия доминанта[64]. «Доверие» к сигналу здесь сравнимо с реакцией на непосредственное появление и опасность агрессии соответствующей птицы (сигнал даже эффективней, рис.17В). Проигрывание «бульканий» подчинённого препятствовало кормлению доминанта, а его предъявление никак не влияло. Сам подчинённый, напротив, не реагировал на запись «бульканий» доминанта, но освобождал кормушку, увидев его вживую (Baker et al., 1991).
А.
Б.
В1.
В2.
Видно, что «бульканье» имеет самостоятельное значение «прогноза» вероятности эффективного подавления сочленов по группе, отдельное от маркёра агрессивности особи (на основе которого, видимо, развился этот сигнал). Дальше, все индивиды в стае гаичек и «зрители», стремящиеся туда включиться[65], вполне «доверяют» информации идеального характера, передаваемой сигналами. «Доверие» проявляется в том, что обе особи, донор и реципиент, выбирают поведение на следующем «шаге» взаимодействия, исходя именно из неё, часто в противоположность «давлению» собственных агрессивных побуждений и «противодавлению» агрессии оппонента.
Примечания:
[60] В выстроенное гнездо самец этого и других видов колюшек одними демонстрациями сперва привлекает самку, затем, в организованном взаимодействии с ней другими демонстрациями побуждает метать икру. Оплодотворив её, самец заботится о потомстве: охраняет гнездо и вентилирует его, усиливая ток воды через него и, соответственно, поступление кислорода к развивающимчся зародышам. У трёхиглой колюшки гнездо шарообразно, у обсуждаемых далее девятииглой Pungitius pungitius и малой южной колюшек P.platygaster напоминает веретенообразную муфту, свитую по спирали.
[61] Интенсивность тех и других в составе «выплеска» прямых действий позволяет определить такую внутреннюю переменную, как мотивация, побуждающую ежей вступать в конфликты друг с другом, а в ответ на демонстрации или действия оппонента не уступать ему, но «повышать планку» как ритуализованной, так и открытой агрессии, стремясь выиграть данный конфликт, тратя на это время, энергию, рискуя получить травму, привлечь внимание хищников и пр. Доля действий противоположного характера (нападения или бегства) в составе выплеска отражает то же самое «соотношение» сил противоположных побуждений (атаковать оппонента, в пределе изгнать его вовсе с участка vs уступать ему, в пределе сдаться и бежать самому), ритуализованно выраженных демонстрациями, для каждого данного момента территориального конфликта. Вообще внутренняя переменная, мотивация особей, создаётся конфликтом побуждений, возникающем при всяком «конфликте интересов» особей во внешнем мире (владелец территории vs захватчик, доминант vs подчинённый в системе агрессивного доминирования, самец vs самка при образовании пары, структурированном так называемой «войной полов» и пр.).
Ритуализованное выражение степени соответствующего конфликта вовне, в виде типологически определённых форм демонстраций — см. разбираемый дальше пример «патрулирования» у ряда видов воробьёв Passer spp. — показывает вероятность разрешения начального конфликта интересов (за ним и производного конфликта мотиваций) в «пользу» данной особи и «против» её оппонента через организованный обмен демонстрациями — коммуникацию. Если оно невозможно из-за отсутствия соответствующих демонстраций в репертуаре вида, как у «непатрулирующих» видов воробьёв или у слётков больших пёстрых дятлов до «созревания» демонстраций ритуализованной угрозы захватчику, то оба конфликта неразрешимы: особи действуют друг против друга чисто физически, сильно стрессируются, рискуют пораниться, тратят время и силы, но безрезультатно. Взаимодействие без коммуникации — просто смертоубийство или, как минимум, сильный риск, снижающийся именно и только при опосредовании прямых действий особей друг относительно друга дифференцированными формами единиц видоспецифического поведения — демонстраций.
По результатам коммуникации одна особь выигрывает, другая проигрывает, т. е. идёт дифференциация ролей, статусов и пр. между исходно «равными» особями, эта созданная коммуникацией разнокачественность оказывает обратное влияние на их поведение при следующем взаимодействии, поддерживая структуру соответствующих отношений в группировке (социальных связей) и одновременно дифференцируя её.
[62] Verrell P.A. The complex courtship of the red-spotted newt// Bull. Chicago Herpetol. Soc. 1987. V.22. №6-7. P.119-120.
[63] См. Доронина Л.О., Володин И.А. Индикаторы эмоционального возбуждения в структуре оборонительных криков большой песчанки (Rhombomys opimus)// IV Всероссийская конф. по поведению животных. Тезисы докл. М.: Товарищество научных изданий КМК, 2007. C.165-166.
[64] И даже символ, поскольку его означаемое (уровень агрессивности) невыводимо из означающего (акустической формы бульканий, представленной на сонаграммах) для наивного наблюдателя, не включённого в эту систему коммуникации: сперва он должен расшифровать эти сигналы.
[65] скажем, бродячие нерезидентные особи (англ. floaters), присутствующие в популяции гаичек наравне с иерархическими группами, занимающими определённые территории в течение всей осени и зимы.
Автор — Владимир Фридман, для «XX2 ВЕКа».
Читайте предыдущие части серии:
20 фактов об инстинктах. Часть 1
20 фактов об инстинктах. Часть 2
20 фактов об инстинктах. Часть 3
20 фактов об инстинктах. Часть 4