
ФАКТ 12
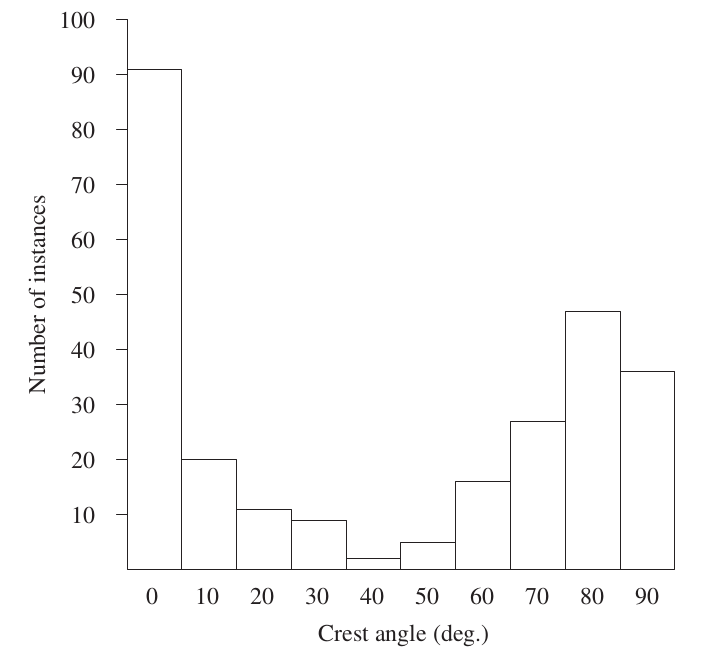
Другой способ обеспечить заметность и одновременно устойчивую распознаваемость форм демонстраций[48] — «квантованность» исполнения образующего движения именно и только при социальной коммуникации, когда выступает релизером, но не в повседневной активности, где это (тело)движение вызвано физическим усилием или неспецифическим возбуждением. Так, у стеллеровой сойки Cyanocitta stelleri движение подъёма хохла в семи различных социальных контекстах (угроза, агрессия, ухаживание, тревога, кроме общего возбуждения и окрикивания хищника) квантовано: самый низкий (0°-10°) или высокий углы подъёма (70°-80° и 90°-90°) намного более часты, чем средние и промежуточные (рис.14).
Другой пример «распада» континуума повседневных движений на элементарные «кванты», моделирующие единичную, двойную, тройную и т.д. «порции» изменения облика животного по каждому из направлений таких изменений (их можно назвать «степени свободы» складывания образов, дальше используемых в демонстрациях) изучено нами на выдвижении крайних рулевых (6-я пара) больших пёстрых дятлов. На них птица не опирается, они делаются заметными (белая кайма, контрастирующая с тёмным хвостом) именно при выдвижении, когда каймы как бы вспыхивают по бокам хвоста (табл.2).
Как и у соек, это движение «квантовано» (рули выдвинуты или наполовину, или полностью, при небольшом числе каких-то ещё значений степени выдвижения)именно и только в территориальных конфликтах, где действует как сильнейшая угроза захватчику (рис.15Б). Именно здесь они демонстрируются подолгу: угрожающий дятел всё время поворачивается спиной и слегка подгибает хвост, как бы «суёт под нос» противнику крайние рулевые. Однако оно полностью континуально («пробегает» все степени выдвижения, от «целиком» до нуля), если вызвано общим возбуждением (видного по раскрытию всего хвоста, в первом случае он закрыт) или физическим усилием, вроде подвешиваний и перепархиваний в ветвях. Другое отличие — вне социальной коммуникации значимо больше нефиксированных, «мимолётных» движений; в территориальных взаимодействиях, даже в паузах, большинство выдвижений крайних рулевых фиксируется у «конечной точки» надолго.
Крайние рулевые D.major имеют рисунок, тип которого (тёмное основание и 2 поперечных полосы) общий с близкими формами, образующими надвид D.major — дятлами белоспинным D.leucotos, средним D.medius, сирийским D.syriacus. Однако лишь у D.major рисунок полос настолько варьирует, что нет двух повторяющихся, он индивидуализирует владельцев территорий в осенне-зимних поселениях, и только здесь выдвижение этих перьев — сигнал территориальной угрозы, наиболее эффективный из выделенных 8 демонстраций охраны территории[49].
В плотных поселениях (>14-15 особей/км²) дятлы жёстко охраняют пространство участка, активно патрулируют по периметру границ (при меньшей плотности охраняем лишь центр и главные «кузницы»). Здесь резиденты направленно подбираются так, что рисунок крайних рулей каждого неповторим и резко отличен от такового соседей, особенно часто вступающих в территориальные конфликты друг с другом; у редко взаимодействующих владельцев участков сходство рисунка значительно выше, оно максимально у птиц, занявших территории в разных концах поселения и не встречающихся. Максимум несходства рисунка также отмечен у брачных партнёров: пары D.major возникают объединением одиночных территорий тех и других во время весеннего возбуждения (Бутьев, Фридман, 2005).
Промежуточная «выдвинутость» рулевых в территориальных конфликтах D.major отмечена много реже, чем у стеллеровой сойки, она всегда мимолётная, не фиксированная, быстро сдвигается к одному из трёх «устойчивых положений» пера (табл.2).
ФАКТ 13
Независимое подтверждение выводов о типологической определённости форм демонстраций, вместе с категориальностью их продуцирования / восприятия во взаимодействиях, где они «работают» как релизеры, получены при постановке первого и второго под оперантный контроль. В обзоре J.D.Pierce (1985) рассмотрена выработка условных рефлексов как инструментальных реакций на разного рода звуки (входящие в число обычных видовых сигналов). Опыты ставили на разных видах приматов, выработав искомый условный рефлекс в 10 случаях. В 9 использовали задачи на дифференцировку, где разными криками надо было обозначать разные раздражители, успешными были 7.
Анализ показывает, что низшие обезьяны способны к контролю звуковой активности, свободно «обозначают» звуками значимые для них вещи или взаимодействия, последнему можно научить и переучить. Возраст, пол и видовая специфика особенно не влияли на результаты этих опытов, а вот подкрепление влияло. Социальное подкрепление было много сильней пищевого, что окончательно подтверждает семантическое кодирование форм демонстраций (Pierce, 1985).
В.Н.Мовчан и А.А.Шибков выработали условный рефлекс избегания ударов током на разные крики видового репертуара, а затем трансформировали их формы, плавно превращая один в другой, анализируя изменения оборонительной реакции на сигналы, всё более непохожие на самих себя:
1) будет она прекращаться быстро и сразу, скачком, при достижении некого порога трансформаций акустической формы, за которой крик Х уже не воспринимается как «тот же самый» или её изменения плавны и постепенны?
2) Будет эта реакция независима от состояния животного? можно ли направленно менять имеющуюся мотивацию проигрыванием сигналов с противоположным значением: скажем, будут ли звуки, связанные с подчинением, тормозить ранее выработанную двигательную реакцию на сигналы угрозы? Контролем в оценке воздействия Х было действие чистых тонов и других криков (У, Z и пр.).
Положительный ответ на 1) говорит о категориальности восприятия криков, составляющих видовой репертуар, на 2) о категориальности продуцирования, проявляющейся вопреки наличию промежуточных звуков между разными криками. Оба «да» были получены для всех изучавшихся землероек, хищников, грызунов и тупай. У кутор Neomys fodiens звуки, связанные с демонстрацией подчинения, эффективно тормозят двигательную реакцию, ранее выработанную на крики угрозы (Мовчан, Шибков, 1987), т.е. действенность связана со спецификой акустической формы, а не стимуляции[50].
Также показана категориальность их восприятия: все вокализации кутор, присущие межиндивидуальным конфликтам, делятся на две категории, резко различных по динамическому спектру. У первых частота спектральных составляющих постоянна, при некотором снижении в конце издавания звука, вторым свойственна синусоподобная частотная модуляция. Физические параметры обоих плавно меняются в зависимости от напряжённости противостояния между зверьками, почему в принципе первые должны нечувствительно трансформироваться во вторые. Но нет, куторы легко разделяют их как разные дискретные сущности, каждая со своим диапазоном варьирования, и выдают рефлекторный ответ лишь на один тип криков. Дифференцирующий их признак формы — характер изменений энергии спектральных составляющих во времени; по этому признаку переход между первыми и вторыми криками скачкообразен, в отличие от изменений звучания, спектра или энергии[51].
Обычные видовые звуки кутор, не связанные с специфическими взаимодействиями, на слух представляют собой континуум переходов от щебета к писку. При помощи той же электрооборонительной методики их разделили на две альтернативные категории, соответствующие разным структурно-ситуативным типам сигналов, переход между ними идёт скачком. Были выявлены границы инвариантного восприятия щебета длительностью 24-30 мс, с энергетически наиболее выраженной частотой 10,4 кГц по отношению к длительности и частоте звуков. Для первого физического параметра это 60 мс, для второго — 6 кГц, если больше — «щебет» сменяется «писком»[52].
Теми же методами у кутор изучили зависимость дифференциальных порогов по частоте от длительности тона частотой 10 кГц. C её уменьшением они увеличиваются: для тонов 500 и 200 мс дифференциальный порог равен в среднем 550 кГц, для тонов 100, 50 и 20 мс - 940, 1570 и 1830 кГц соответственно. Затем установили роль спектральных характеристик в распознавании куторами ещё одного видоспецифического сигнала, короткого «джжж». Его надёжность распознавания обеспечивается наличием в модели сигнала как минимум двух спектральных максимумов; исключение одного из них, высоко- или низкочастотного, резко снижает точность.
У орешниковой сони Muscardinus avellanarius структурный анализ вокализаций сперва выделил три модификации визгов, сопровождающих оборонительное поведение: пронзительный визг, стрекочущий визг и визг самки при спаривании. Они различаются более-менее надёжно по ряду параметров звучания: частоте энергетически наиболее выраженной спектральной составляющей, длительности отдельных импульсов, характеру динамического спектра и по временнόй организации[53]. Различия чётки, пока животное спокойно. Рост возбуждения или страха заставляет вокализации «плыть»; если анализировать только сонограммы, их акустические формы будут «смазаны» или связаны переходами, но выработка реакции избегания с этими криками как условными раздражителями показывает точное соответствие между формальной классификацией визгов и их восприятием сонями.
Для различения визгов с отделением друг от друга сони используют совокупность трёх признаков: широкополосность гармонического звука, наличие/отсутствие повышающейся частотной модуляции, расположение первой гармоники в области частот ниже 10 кГц. Разные типы визгов заданы как разные комбинации состояний трёх ключевых признаков, устойчивые к изменениям уровня мотивации. Поэтому в любой момент зверёк может определить, есть сигнал, или нет, и какой.
Дальше исследовали восприятие сонями стрёкота, отличающегося ритмически организованной быстрой последовательностью коротких частотно-модулированных звуков. Та же методика показывает, что искажение временных параметров стрёкота снижает уровень положительных реакций лишь незначительно; сильное снижение отмечено лишь при предъявлении криков, точно соответствующих стрёкоту по длительности импульсов, интервалу между импульсами и т.п., но с полностью изменённой формой динамического спектра сигнала. Важнейший структурный признак, позволяющий распознать «нужный» стрёкот среди сходных звуков — скорость частотной модуляции; кричащие сони её удерживают в рамках, обеспечивающих уверенное распознавание[54]. Иными словами, стрёкот распознаётся по специфической форме сигнала, а не по интенсивности акустической стимуляции. Если б крики действовали только как стимулы, одни и те же характеристики их акустической формы отвечали бы за распознавание и за действие на реципиента, однако они строго разделены — крик распознаётся по одним характеристикам, а воздействие обеспечивают другие.
Дальше использованием той же методики показали, что сони не только воспринимают разные видовые крики как отдельные дискретные сущности, но и классифицируют их на основе использования как сигналов, т. е. по семантической близости в неком ряду (называемому обычно сигнальным: он соответствует какому-то определённому типу проблемных ситуаций, разрешаемых во взаимодействиях, и типу компаньонов — потенциальных участников коммуникации). При классификации близость семантики издаваемых звуков, интерпретируемых как «один из сигналов ряда», даже важнее близости их акустической структуры[56].
В опытах с орешниковыми и садовыми сонями Eliomys quercinus показана способность зверьков к тонкой дифференциации звуков. Они разделяют звуки, отличные по частоте на 8% у первой и 13% у второй, по длительности на 13%, причём всё это при невозможности непосредственного сличения. Следовательно, существуют нервные модели всех значимых видовых криков, с которыми происходит сравнение[56].
Категориальность восприятия и «семантичность» классификации сигналов зафиксирована и в вокализациях домашних кошек Felis catus, соответствующих начальной фазе конфликта. Ей присущи звуки разной длительности (0,4-6,5 с) и спектрального состава (частота 1-й гармоники 0,15-1,75 кГц), на слух воспринимающиеся как вои, вопли, вскрики, мяуканья, урчание и шипение. При помощи той же методики изучали изменчивость значений физических параметров данных звуков, и особенности их восприятия кошками, классификацию сигналов, значимую для самих кошек, и идентификацию ими разных вокализаций на принадлежность к тому или иному типу, выделенному по данным структурного анализа. Для кошек воспринимаемый на слух континуум криков делится на две контрастирующие отдельности: мяуканье-высокочастотные вои и низкочастотные вои разной длительности. Когда котам предъявляли ранее незнакомые звуки первого типа, условная реакция отмечена в 68-98% случаев, на звуки второго типа её нет вовсе[57].
Подытоживая результаты экспериментов по проигрыванию разным видам млекопитающих естественных и «искажённых» сигналов с целью выделения структурно-ситуативных типов звуков, имеющих «значение» для самих особей, В. Н. Мовчан показывает, что «размер» выделенной категории акустических сигналов и чёткость её отделения от других категорий сильно зависят от показателя реакции, выбранного для эксперимента. В опытах с плавным искажением формы сигналов, «превращающим» их друг в друга, выделенность акустических демонстраций и точность дифференцирования их между собой сильно зависят от ожидаемых реакций животных в опыте – нужно ли им «действовать специфическим образом» или «волноваться». В первом случае формы демонстраций дискретизуются много точнее, граница между ними в последовательности проводится чётче и однозначней[58].
Частота сердечных сокращений, обычно используемая в анализе действенности акустических сигналов, мало избирательна и слабо надёжна. Видимо, она отражает наиболее общий уровень классификации акустических сигналов, основанный на оценке важности для жизнеобеспечения. Наибольшие избирательность реагирования на разные вокализации, как и максимум точности различения их в опытах с «трансформацией формы», выявлены при использовании выработанной на них условно-рефлекторной реакции[59]. Т.е., как было сказано выше, специфика форм или образов демонстраций, развёртывающихся во внешнем мире, животным существенней изменений их внутренних состояний, и это важнейшая характеристика осуществления инстинкта.
Примечания:
[48] Т. е. устойчиво и точно дифференцировать их друг от друга или от «фона» в виде континуума прямых действий или экспрессивных реакций.
[49] Но не у близких форм, где отсутствует и столь жёсткая одиночная территориальность осенью и зимой. У белоспинного дятла участки не охраняются, у сирийского пара сохраняется на зиму, птицы охраняют лишь центр участка.
[50] Это независимое подтверждение вывода, что с точки зрения семиотики ритуализированные демонстрации представляют собой произвольный знак, как и другие классические релизеры.
[51] См. Мовчан В.Н., Шибков А.А. Опознание землеройками внутривидовых коммуникационных сигналов и их синтезированных моделей // Вестник ЛГУ. 1987. №1. С.67-84.; Шибков А.А., Мовчан В.Н., Соболевский С.А. Категориальный характер восприятия землеройками сем. Soricidae звуковых сигналов, сопровождающих конфликтные взаимодействия между животными// Нервная система. 1990. №29. С.61-74.
[52] Соболевский С.А., Мовчан В.Н., Шибков А.А. Восприятие и анализ коротких звуков куторой обыкновенной (Neomys fodiens)// Ред. журн. "Вестник ЛГУ. Биол.". Л., 1988. 10 с. Рукопись деп. в ВИНИТИ 04.04.88, №2536-888.
[53] Мовчан В.Н., Коротецкова Л.В. Опознавание орешниковой соней Muscardinus avellanarius (Rodentia; Gliridae) акустических сигналов, сопровождающих агонистическое поведение// Зоол. ж. 1987а. Т.66. №9. С.1363-1374.
[54] Мовчан В.Н., Коротецкова Л.В. Восприятие соней Muscardinus avellanarius последовательностей коротких частотно-модулированных звуков// Журнал эволюционной физиологии и биохимии. 1987б. Т.23. №5. С.635-641.
[55] Коротецкова Л.В., Мовчан В.Н. Классификация видоспецифических звуковых коммуникационных сигналов грызунами семейства соневых Gliridae // Нервная система. 1990. №29. С.75-88.
[56] Мовчан В.Н., Коротецкова Л.В. Оценка частотных и временных параметров акустических сигналов представителями семейства сонь (Gliridae) // Вестник ЛГУ. 1985. №24. С.84-87.
[57] Мовчан В.Н., Чернышёва Т.В. О дискретном восприятии домашней кошкой сигналов, сопровождающих акустическое поведение// Коммуникативные механизмы регулирования популяционной структуры у млекопитающих. М.: Наука, 1988. С.87-89.
[58] Мовчан В.Н. Экспериментальный анализ критериев оценки восприятия млекопитающими акустических коммуникационных сигналов// Мат-лы V Съезда Всес. теориол. Общ-ва АН СССР, Москва, 29 янв. – 2 февр. 1990. М.: 1990. С.36-37.
[59] См. В.М.Мовчан, 1990. op.cit.
Автор — Владимир Фридман, для «XX2 ВЕКа».
Читайте предыдущие части серии:
20 фактов об инстинктах. Часть 1
20 фактов об инстинктах. Часть 2
20 фактов об инстинктах. Часть 3
20 фактов об инстинктах. Часть 4
20 фактов об инстинктах. Часть 5