1 Сахар, искусственные подсластители и ожирение
Сахар вошел в пищевую цепочку в конце 1960-х годов и заменил жиры, чтобы скрыть горечь и сделать пищу более вкусной (Bakke et al., 2018; Moss, 2013).
В 1970-х годах стал очевиден сдвиг в сторону более подслащенных сахаром напитков (Wolf et al., 2008). Наши ранние предки получали сахар либо из фруктов, ограниченных сезонами, либо из меда, защищенного пчелами. За последние полвека потребление сахара утроилось во всем мире, отчасти из-за скрытого использования добавленных сахаров в обработанных пищевых продуктах (Lustig et al., 2012). Первый искусственный подсластитель (сахарин) был представлен в 1879 году с низкими производственными затратами во военное время, что увеличило его популярность (Weihrauch and Diehl, 2004). В 1950-х годах, когда сахар стал легкодоступным, использование подсластителей перешло на так называемые «диетические продукты» с низкой калорийностью.
Развитие ожирения зависит как от гедонистического, сладкого вкуса пищи в сочетании с отрицательными эмоциональными свойствами потребления пищи (Meye и Adan, 2014; Yu и Kim, 2012; Berthoud и др., 2017). Гедонистические реакции на 10% раствор сахарозы (и даже на 3,4%) были протестированы и оказались значительно выше у подростков крыс по сравнению со взрослыми крысами (Wilmouth и Spear, 2009).
Сахар и эмоции
Было показано, что эмоциональное питание возникает из-за желания смягчить последствия стресса (Van Strien и др., 1986), а стресс частично регулируется гипоталамо-гипофизарно-надпочечниковой (HPA) осью.
Интересно, что активность оси HPA, как было показано, снижается за счет потребления продуктов, содержащих сахар (Ulrich-Lai et al., 2011).
После потребления выделяются гормоны, которые уменьшают чувство стресса, что также увеличивает желание пищи для комфорта, тем самым закрепляя эмоциональные привычки питания (Ulrich-Lai et al., 2011; Ursano et al., 2009).
Цель этого обзора — обобщить исследования, которые изучают, как потребление сахара приводит к изменениям в нейробиологической функции мозга, которая изменяет эмоциональное состояние и последующее поведение.
Мы рассмотрим результаты исследований на пересечении между потреблением сахарозы и изменениями в выполнении задач со стрессовым или эмоциональным компонентом и рассмотрим нейробиологические и нейрохимические механизмы, вовлеченные в зависимость, стресс, страх, тревогу и депрессию, чтобы определить, существуют ли перекрывающиеся нейронные механизмы.
Наконец, мы определим, существуют ли новые фармакотерапевтические средства и/или вмешательства, нацеленные на эти мозговые контуры или нейрохимические пути для улучшения текущих подходов к лечению ожирения.2. Общие нейронные пути для потребления сахарозы, зависимости, эмоций и ожирения Зависимость характеризуется трудностью контроля привычного поведения даже перед лицом негативных последствий (Lindgren et al., 2018).Ранние исследования зависимости были сосредоточены на наркотиках, вызывающих злоупотребление, таких как алкоголь, морфин и никотин.
С тех пор это было расширено, чтобы включить азартные игры, еду и, в последнее время, потребление сахара (Comings et al., 2001; Weeks, 1962; Jellinek, 1952; Benowitz, 1988; Gearhardt et al., 2009; Klenowski et al., 2016; Benton, 2010).
Зависимость от веществ A. Jacques, et al.злоупотребление зависит от связывания препарата с определенными белковыми мишенями, что вызывает определенные физиологические и поведенческие реакции, уникальные для этого препарата (Lee and Messing, 2008).
Психоактивные препараты обычно вызывают приятные ощущения, которые приводят к повторному использованию и, в зависимости от генетической восприимчивости, факторов окружающей среды и последующей зависимости (Lee and Messing, 2008).
Стресс давно ассоциируется как с мотивацией к использованию полезных веществ, так и с результатом неполучения этих веществ (для обзора см. (Sinha, 2008)).
Негативные симптомы, вызванные отменой, являются общими для всех форм наркотических веществ, включая очень вкусную пищу.
К ним относятся длительная сенсибилизация к выбранному веществу и ассоциативное обучение, когда сигналы окружающей среды становятся связанными с удовольствием, получаемым от вещества (Hebebrand et al., 2014).
Эти ассоциативные воспоминания, в сочетании с сильными тягами, увеличивают частоту рецидивов даже после длительного воздержания.
Эти общие черты могут быть обусловлены тем, что вещества, вызывающие зависимость, используют те же схемы в мезокортиколимбической системе мозга (рис. 1) (Volkow et al., 2004; Koob and Le Moal, 2001; Baron et al., 2018).
Потребление пищи необходимо для регулирования гомеостатического энергетического баланса.
Однако люди также потребляют пищу для удовольствия или комфорта.
Гедонистическое желание вкусной пищи считается связанным с вознаграждением и может привести к неадаптивной или отрицательной нейропластичности, которая может пересиливать гомеостатическую регуляцию и приводить к перееданию (Kenny, 2011). Вознаграждение предоставляется через стимулы, которые вызывают приятные или приятные переживания, в отличие от зависимости, которая включает компульсивное и иногда болезненное поведение.
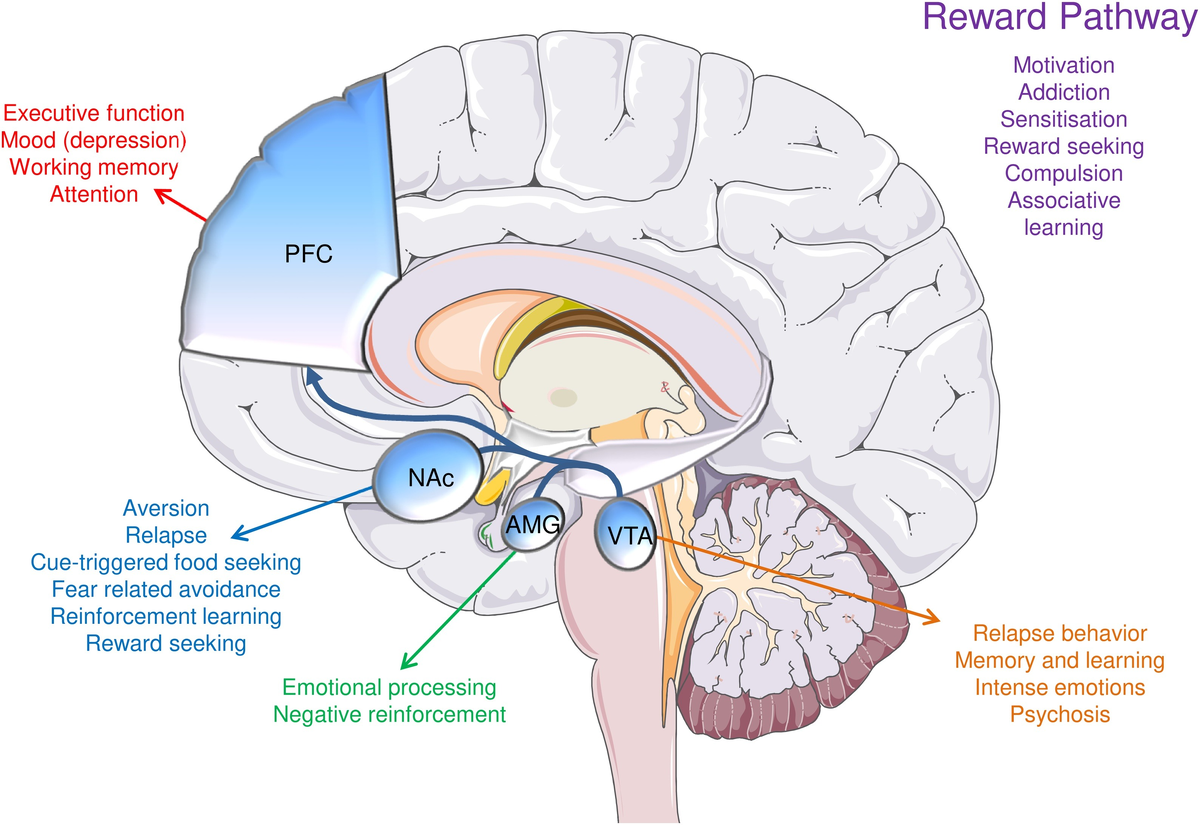

Рис. 2. Регуляция пищевого поведения и потребления пищи центральными и периферическими гормонами и пептидами, регулирующими аппетит. Гипоталамические орексигенные и анорексигенные пути и их регуляция пептидами NPY/AgRP и POMC/CART соответственно изображены на правой боковой панели.
Также изображены периферические гормоны и пептиды, стимулирующие аппетит (зеленый) и ингибирующие (красный), пересекающие гематоэнцефалический барьер (ГЭБ), а также органы, из которых они происходят.
Влияние этих гормонов/пептидов на стимуляцию или ингибирование орексигенных (пунктирные зеленые стрелки) и анорексигенных (пунктирные красные стрелки) обозначено как (+) или (-) соответственно.
Боковые гипоталамические орексиновые нейроны получают входные данные о потреблении пищи от дугообразного ядра гипоталамуса через эндогенные антагонисты меланокортиновых рецепторов; нейропептид Y (NPY) и нейроны, экспрессирующие агути-родственный пептид (AgRP).
Кроме того, метаболический гомеостаз воспринимается латеральным гипоталамусом через окружающие уровни глюкозы, грелина и лептина, и это управляет поведением поиска пищи (Yamanaka et al., 2000).
Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors: Cell
Гипоталамические орексинергические и анорексинергические пути регулируются пептидами NPY/AgRP и POMC/CART и по-разному подвергаются влиянию потребления сахара (Murray et al., 2016). Орексигенный путь находится под влиянием двух специфических популяций нейронов в дугообразном ядре, первая популяция экспрессирует NPY и AgRP и стимулирует потребление пищи (Benoit et al., 2008; Yamanaka et al., 2000).
Изменения как в NPY, так и в AgRP были связаны с тем, что крысы потребляли больше пищи после того, как им давали 30% раствор сахарозы (Gaysinskaya et al., 2011).
Снижение экспрессии этих пептидов было замечено после потребления сахарозы, которая затем увеличивалась через тридцать-шестьдесят минут, до кормления (Gaysinskaya et al., 2011). Связь между p53 (геном, кодирующим белок, участвующий в регуляции клеточного цикла) в нейронах AgRP (которые регулируют аппетит, вызванный грелином) и ожирением была продемонстрирована на мышах, причем его сверхэкспрессия приводила к чрезмерной потере веса (Quiñones et al., 2018).
В нормальных условиях недостаток доступных питательных веществ побуждает нейроны AgRP инициировать пищевое поведение. Было показано, что у мышей желание есть обязательно включает активацию нейронов AgRP, когда пища была обогащена сахаром и жиром (Denis et al., 2015).
В этом случае сигнализация дофамина инициирует желание есть, и пищевое поведение становится обусловленным вознаграждением через нейронные цепи, вовлеченные в эмоции, в отличие от орексигенного пути, вызванного метаболической потребностью.
По-видимому, это имеет место в случае еды для комфорта (Denis et al., 2015). Дальнейшие исследования напрямую связывают потребление сахарозы с орексигенным путем.
В двух отдельных исследованиях было обнаружено, что потребление сахарозы увеличивается после инфузии NPY в боковой желудочек или введения AgRP в оболочку NAc (Badia-Elder et al., 2003; Pandit et al., 2015). Однако эффекты введения AgRP в оболочку NAc останавливаются предварительной обработкой α-флупентиксолом, неселективным антагонистом дофаминовых рецепторов (Pandit et al., 2015). Кроме того, хемогенная и оптогенетическая манипуляция активностью нейронов AgRP модулирует эмоциональную валентность кормления. В частности, было показано, что нейроны AgRP дугообразного ядра регулируют эмоциональные аспекты кормления, связанные с тревогой, поведением, подобным страху, и агрессией (Sweeney и Yang, 2017; Burnett и др., 2016; Joly-Amado и др., 2014; Padilla и др., 2016).
Кроме того, серотонинергическая система показана в регуляции гедонистического питания, при этом увеличение серотонина вызывает снижение потребления пищи, а снижение серотонина увеличивает мотивацию к еде (Halford et al., 2010).
Предполагаемый механизм действия серотонина в поведении питания осуществляется через нейроны AgRP и POMC в дугообразном ядре (Sohn et al., 2013).
Полиморфная область, связанная с транспортером серотонина (5-HTTLPR), регулирует уязвимость к стрессу, которая увеличивается в случаях патологического страха, и влияет на потребление энергии, что предполагает ее роль в переедании, связанном со стрессом (Capello and Markus, 2014).
Нейронные адаптации, возникающие в результате сахарной зависимости у грызунов, включают изменения в связывании дофамина и опиоидных рецепторов в мезолимбической коре, измененную экспрессию мРНК энцефалина и измененное высвобождение дофамина и ацетилхолина NAc (Avena et al., 2008).
Лептин и грелин
Существует несколько альтернативных нейронных популяций, путей и ядер мозга, которые управляют поведением переедания и потреблением сахарозы. Наряду с нейронами гипоталамуса, продуцирующими орексин, нейроны MCH и клетки рецепторов лептина, выступающие из латерального гипоталамуса в VTA, также влияют на гедонистическое питание и поведение поиска вознаграждения (Bocarsly, 2018).
Высокое потребление фруктозы приводит к более низким уровням инсулина, снижению уровня лептина и повышению концентрации грелина по сравнению с пищей с высоким содержанием глюкозы (Teff et al., 2004).
Изменение уровней этих гормонов сытости (которые приводят к чувству «сытости» после приема пищи) может быть предшественником переедания. Эти результаты совпадают с результатами, полученными после интрацеребровентрикулярной инъекции концентрированной фруктозы или глюкозы в гипоталамус крыс, подвергшихся воздействию 2-дезокси-D-глюкозы (2DG) (Miller et al., 2002).
DG является аналогом глюкозы, который не может метаболизироваться и, как известно, вызывает повышенное потребление пищи у крыс, вмешиваясь в процесс гликолиза (Fiorentini and Müller, 1975). Крысы, которым вводили инъекции фруктозы, показали повышенное потребление пищи как в присутствии, так и в отсутствие 2DG, где глюкоза подавляла вызванное 2DG потребление пищи, что предполагает роль глюкорецепторов мозга в контроле потребления пищи (Miller et al., 2002).
Глюкорецепторы контролируют уровень глюкозы в крови и расположены в гипоталамусе.
Было показано, что они стимулируются низким уровнем сахара в крови, что приводит к увеличению пищевого поведения (Ritter et al., 1981).
Дофаминергические нейроны, проецирующиеся от VTA к NAc, ингибируются лептином и инсулином и стимулируются грелином (Palmiter, 2007).
Было показано, что чрезмерное потребление сахара увеличивает связывание рецептора дофамина D1 в ядре и оболочке NAc, уменьшает связывание дофамина D2 в дорсальном полосатом теле и увеличивает связывание с транспортерами дофамина в среднем мозге (Colantuoni et al., 2001; Bello et al., 2002).
Увеличение дофамина задерживает высвобождение ацетилхолина во время кормления, что откладывает насыщение и прокладывает путь к перееданию (Hoebel et al., 2009).
Альтернативно, связывание опиоидных рецепторов (mu-1) увеличивалось в оболочке NAc, гиппокампе, голубом пятне и поясной коре после того, как крысам разрешалось периодически употреблять 25% раствор глюкозы (Colantuoni et al., 2001).
В совокупности эти результаты свидетельствуют о том, что чрезмерное потребление сахара может сенсибилизировать дофаминовые D1 и опиоидные рецепторы mu-1 аналогично наркотикам, вызывающим зависимость.
В таблице 2 представлены исследования, изучающие влияние потребления сахара на области мозга, молекулярные механизмы и поведенческие изменения, связанные с орексигенным путем.
Компульсивный поиск сахарозы
С 1980-х годов было известно, что нейроны из латерального гипоталамуса кодируют сигналы, связанные с вознаграждением (Nakamura et al., 1987), а также как пищевое (Schwartzbaum, 1988), так и питьевое (Tabuchi et al., 2002) поведение.
В 2015 году была предпринята попытка определить нейронные цепи, специфичные для компульсивного поиска сахарозы (Nieh et al., 2015).
Фотоингибирование использовалось для того, чтобы показать, что избирательный путь от латерального гипоталамуса к VTA снижает компульсивный поиск сахарозы у мышей, не влияя на нормальное пищевое поведение (Nieh et al., 2015).
Исследование выявило двунаправленную цепь как ингибирующих, так и возбуждающих проекций от латерального гипоталамуса к дофаминергическим и ГАМКергическим нейронам в VTA (Nieh et al., 2015).
Возможно, что эта цепь увеличивает фокус и интенсивность мотивации сахарозы, выделяя ее как потенциальную терапевтическую цель для компульсивного переедания сахара.
Поскольку ожирение возникает из-за избыточного потребления калорий, стимулирующая ценность потребляемой пищи должна рассматриваться параллельно с насыщением голода. Рецепторы 5-гидрокситриптамина 2C (5-HT2CR) играют роль в стимулирующей мотивации через цепи питания гипоталамуса-VTA и поэтому считаются целью для лечения ожирения (Valencia-Torres et al., 2017).
Лоркасерин, агонист 5-HT2CR, вводился мышам и, как было показано, снижал как стандартное потребление пищи, так и желание шоколадных шариков с соответствующим увеличением экспрессии c-fos в ГАМКергических нейронах VTA 5-HT2CR (не дофаминергических нейронах) (Valencia-Torres et al., 2017), подтверждая их роль в ингибировании мотивационного поведения. Аналогичные результаты были получены при наблюдении за употреблением сахарозы, когда было обнаружено, что лептин, вводимый в VTA, снижает потребление пищи, а подавление рецепторов лептина увеличивает поиск сахарозы (Clifton, 2017).
Интересно, что посредничество орексигенного пути между VTA и NAc может влиять на поведенческие изменения в сторону поиска вознаграждения за наркотики, вызывающие зависимость, включая поведение отмены и рецидива (Kastin, 2013; Carr and Kalivas, 2006).
Снижение уровня дофамина и одновременное повышение уровня ацетилхолина связаны с поведенческими признаками отмены наркотиков, вызывающих зависимость (Rada et al., 2001; Rada and Hoebel, 2005).
Крысы, которым предоставлялся прерывистый доступ к сахару, демонстрируют схожие дисбалансы в инициировании отмены после предоставления антагониста опиоидных рецепторов налоксона (Colantuoni et al., 2002).
Животное будет жаждать вещества, вызывающего зависимость, которого оно было лишено, о чем свидетельствует повышенная оперантная реакция даже без вознаграждения и повышенная реакция на сопутствующие сигналы о наркотиках (Bienkowski et al., 2004; Lu et al., 2004).
Крысы, лишенные сахара после чрезмерного потребления глюкозы (т. е. которым разрешалось 25% глюкозы в течение 30 минут в день в течение 28 дней и доступ к глюкозе в их домашних клетках в течение дополнительных 11,5 часов в день), реагировали значительно сильнее в оперантных камерах по сравнению с контрольной группой, что свидетельствует о том, что сенсибилизация дофаминергической системы и ассоциативное обучение приводят к повышению мотивации к поиску сахара (Avena et al., 2005).
Нейронная адаптация, необходимая для того, чтобы вызвать изменение поведения, кажется, сохраняется долго, поскольку потребление сахара прекращается за 2 недели до тестирования.
Аналогичные результаты были получены в исследованиях потребления алкоголя на крысах (Heyser et al., 1997).
Потребление сахарозы и гипоталамо-гипофизарно-надпочечниковая ось
Длительный стресс, в зависимости от его тяжести, по-видимому, коррелирует с предпочтением продуктов с высоким содержанием сахара, что предполагает его вклад в прогрессирование ожирения.
Хронический стресс может развиться из-за накопления физических (травматических), химических (диетических), физиологических (болезненных), психологических (страх) или социальных (образ жизни) стрессоров (Powers and Howley, 2007).
Длительные периоды стресса приводят к гиперактивации оси гипоталамус-гипофиз-надпочечники (HPA), системы реакции млекопитающих на стресс, включающей эндокринную и центральную нервную системы (Smith and Vale, 2006).
ОСЬ ГИПОТАЛАМУС-ГИПОФИЗ-НАДПОЧЕЧНИК
Ось приводит к повышенному высвобождению кортикостероидов (стероидных гормонов, вырабатываемых в коре надпочечников), см. рис. 3 (Smith and Vale, 2006; BjÖrntorp, 2000).
Активация оси приводит к повышению уровня адренокортикотропного гормона (АКТГ) и повышению уровня глюкокортикоидов, которые влияют на использование запасов энергии (Ulrich-Lai et al., 2010).
Figure 1. Schematic representation of the hypothalamic-pituitary-adrenal (HPA) axis. Hypophysiotropic neurons localized in the paraventricular nucleus (PVN) of the hypothalamus synthesize corticotropin-releasing factor (CRF) and vasopressin (AVP). In response to stress, CRF is released into hypophysial portal vessels that access the anterior pituitary gland. Binding of CRF to the CRF type 1 receptor (CRFR1) on pituitary corticotropes activates cyclic adenosine monophosphate (cAMP) pathway events that induce the release of adrenocorticotropic hormone (ACTH) into the systemic circulation. In the presence of CRF, AVP elicits synergistic effects on ACTH release that are mediated through the vasopressin V1b receptor.
Circulating ACTH binds to the melanocortin type 2 receptor (MC2-R) in the adrenal cortex where it stimulates glucocorticoid synthesis and secretion into the systemic circulation.
Glucocorticoids regulate physiological events and inhibit further HPA axis activation (red lines) through intracellular receptors that are widely distributed throughout the brain and peripheral tissues. IP3, inositol triphosphate; DAG, diacylglycerol .
Это имеет особое значение, поскольку люди регулярно сообщают о выборе потребления сладкой пищи (более калорийной) из-за ее способности улучшать их настроение и снимать негативные эмоциональные состояния (Smith and Vale, 2006).
Figure 2. Depiction of the major brain regions and neurotransmitter groups that supply afferent innervation to the medial parvocellular zone of the paraventricular nucleus (PVN). Cell groups of the nucleus of the solitary tract (NTS) and ventral medulla (C1) relay visceral information to the PVN though noradrenergic (NE), adrenergic (Epi), and glucagon-like peptide 1(GLP-1) containing neurons. Hypothalamic nuclei (HYPO) encode information from endocrine systems and send mainly γ-aminobutyric acid (GABA)-ergic (GABA) projections to the PVN. Cell groups of the lamina terminalis relay information concerning the osmotic composition of blood to the PVN through glutamatergic (Glu) and angiotensinergic (Ang) neurons. Limbic structures including the hippocampus, prefrontal cortex, and the amygdala contribute to the regulation of PVN neurons through intermediary neurons of the bed nucleus of the stria terminalis (BNST). PIT, pituitary. Adapted from reference 20: Sawchenko PE, Imaki T, Potter E, Kovacs K, Imaki J, Vale W. The functional neuroanatomy of corticotropin-releasing factor. Gba Found Symp. 1993;172:5-21; discussion 21-29. Copyright © John Wiley and Sons 1993.
В наблюдении за крысами, подвергающимися острому стрессу, предоставление раствора сахарозы значительно снизило уровни секреции как АКТГ, так и кортикостерона (Ulrich-Lai et al., 2010).
Кроме того, крысы, которых кормили подсластителем (сахарином), также показали более низкие реакции оси HPA на острый стресс, что позволяет предположить, что гедонистическая природа сахара может быть ответственна за сниженную реакцию на стресс (Ulrich-Lai et al., 2010).
Последствия потребления сахарной сукросы и апартемии в подслащенных напитках на региональную мозговую активность.
Прединтервенционные изображения мозга, показывающие эффекты стрессового задания (MIST) на посещении прединтерварии, обвалились по всем участникам из обеих групп вмешательства. Стресс-задача привела к значительной (P < .01) односторонней (левая панель) деактивации в миндалине ( 20, 4, 16), гиппокампе ( 22, 16, 22) и передней cingulata коре (BA10; 4, 52, 12). В, повторно-меры ANCOVA показали значительную группу лечения по посещению взаимодействия [F (1, 15) = 6.8, P = 020; размер эффекта (η 2p) = 0,15]. По сравнению с прединтервенционной проверкой, значительно большая гиппокамповая активность, вызванная MIST, была обнаружена в группе сахароза, но не апартамы, на послеоперационной проверке. Изображение мозга в панели С показывает чистый эффект потребления сахарозы на увеличение довмешательства и апартатома...
Напротив, исследование с участием девятнадцати женщин показало, что потребление сахарозы, но не подсластителя (аспартама), привело к снижению кортизола, вызванного стрессом (Tryon et al., 2015).
ПОДСЛАСТИТЕЛИ
https://pmc.ncbi.nlm.nih.gov/articles/PMC4454811/
Сахароза, а не аспартам, также вызывала большую активность в левом гиппокампе (Tryon et al., 2015), что позволяет предположить, что отрицательная обратная связь может способствовать формированию тенденции к потреблению сахара у людей, сталкивающихся со стрессом.
В следующем разделе рассматривается сложная взаимосвязь между осью HPA, потреблением пищи, запасами энергии и хроническим стрессом.
Любой выявленный острый стрессор может вызвать вовлечение оси HPA и привести к эмоциональной реакции, вызывающей страх, беспокойство или защитное поведение, призванное максимизировать шансы на выживание (Ulrich-Lai et al., 2010).
Связь между стрессом, уровнем кортизола и потреблением сахара далека от понимания, однако, по-видимому, диеты с высоким содержанием сахара играют важную роль в регуляции оси гипоталамус-гипофиз-надпочечник и перееданию, связанном со стрессом.
При употреблении в пищу продукты с высоким содержанием сахара высвобождают нейропептиды, повышая настроение и усиливая желание или избирательное предпочтение к большему количеству высококалорийной пищи (Ulrich-Lai et al., 2011).
Было показано, что сахароза и подсластитель сахарин ослабляют реакцию оси HPA, и этот эффект, возможно, является причиной того, что очень вкусные продукты потребляются для смягчения неприятных эмоций (Ulrich-Lai et al., 2011).
Потребление сахара регулирует стрессоподобное поведение через ось HPA, однако точные механизмы действия еще предстоит выяснить.
Напротив, примеры хронического стресса показывают повышенные уровни глюкокортикоидов с повышенным риском развития заболеваний, связанных со стрессом, таких как депрессия.
Длительный стресс, по-видимому, изменяет функцию мозга за счет изменений в отрицательных обратных связях от энергетических запасов и глюкокортикоидной модуляции нейронных цепей (для обзора см. (Dallman et al., 2003)).
Модели, представляющие острую и хроническую реакцию GC на функцию в оси HPA. Канонические эффекты происходят быстро, в течение нескольких минут или часов после стресса; GC действуют непосредственно на мозг и гипофизу, вероятно, через негеномические механизмы. Новая модель требует 24 часа после возбуждения GC в концентрацию стресса. Тогда, прямое действие ГК на мозг стимулирует, и отрицательная обратная ингибиция функции в оси HPA является следствием метаболических эффектов GC увеличение накопленных энергии брюшных.
Поскольку уровни глюкокортикоидов строго контролируются осью HPA, одним из направлений исследований было изучение уровней экспрессии мРНК 11βHSD-1 (медиатора метаболизма глюкокортикоидов в печени) в случаях стресса в раннем возрасте (Maniam et al., 2015).
Заинтересовавшись ролью потребления сахара в гомеостазе липидов, исследователи обнаружили 53%-ное увеличение транскрипционных уровней 11βHSD-1 у животных, которым разрешалось хроническое потребление сахара (25% раствор сахарозы), в отличие от контрольной группы, выявившей корреляцию между потреблением сахарозы, стрессом и повышенным метаболизмом глюкокортикоидов (Maniam et al., 2015).
Кроме того, когда люди подвергаются стрессу в молодом возрасте, у них повышается вероятность развития тревожности и депрессии, связанных с нарушением регуляции оси HPA (Heim and Nemeroff, 2001).
Повышенный уровень кортизола (результирующий продукт оси HPA у людей) также положительно коррелировал с более высокими отложениями висцерального жира и резистентностью к инсулину (Misra et al., 2008).
Аналогично высокий уровень кортизола (на 22% выше) был зарегистрирован у подростков с избыточным весом/ожирением после 2 или более подслащенных сахаром напитков в день, что предполагает значительное увеличение гормона стресса не только из-за отложений висцерального жира, но и из-за повышенного потребления сахара (Shearrer et al., 2016).
Один из предполагаемых механизмов может включать морфологическое изменение надпочечников, как показано при потреблении напитков с высоким содержанием сахара, что увеличивает риск метаболических синдромов, таких как диабет, и приводит к дисфункции надпочечников (Díaz-Aguila et al., 2016).
Молодые взрослые крысы, которым в течение 12 недель давали 30% раствор сахарозы, показали увеличение отложений висцерального жира и резистентность к инсулину (Díaz-Aguila et al., 2016).
Интересно, что надпочечники показали гистоморфологические изменения в корковом и мозговом веществе надпочечников из-за потребления сахарозы, что указывает на гиперплазию и признаки метаболического синдрома (Díaz-Aguila et al., 2016).
Нейроадаптация в процессе развития
У грызунов пренатальный стресс может снижать уровни глюкокортикоидных и минералокортикоидных рецепторов в гиппокампе, тем самым уменьшая доступность рецепторов для ингибирования обратной связи кортикостерона, что может объяснить, почему новые стрессоры вызывают повышенную и более длительную реакцию кортикостерона (Henry et al., 1994).
Беременные крысы подвергались стрессу ограничения в течение третьей недели беременности (Henry et al., 1994).
Были исследованы ось HPA и гиппокампальные кортикостероидные рецепторы у самцов.
Было обнаружено, что плазменный кортикостерон был значительно выше у крыс, подвергшихся пренатальному стрессу, по сравнению с контрольной группой, а подтипы рецепторов (гиппокампальные кортикостероидные рецепторы типа I и типа II) были снижены в гиппокампе, что предполагает долгосрочные изменения в оси HPA, которые могут возникнуть после пренатального стресса (Henry et al., 1994).
Многочисленные исследования изучали материнский стресс и кортикостероиды, для всестороннего обзора см. (Welberg and Seckl, 2001).
Меньше исследований включают эти нейроадаптации и диету, поэтому более прямое исследование высокого потребления сахара у матерей может быть значимым в изучении детского ожирения.
Кроме того, существуют доказательства предварительного программирования гиперактивации оси HPA в пренатальные периоды (Welberg и Seckl, 2001).
У подростков, которые, как считается, подвержены более высокому риску ожирения (оценивая индекс массы тела их родителей), более высокий функциональный сигнал магнитно-резонансной томографии (фМРТ) наблюдался в областях мозга, связанных с обучением, обработкой и мотивацией вознаграждения после потребления молочных коктейлей с высоким содержанием сахара по сравнению с молочными коктейлями с высоким содержанием жира (Shearrer et al., 2018a)
Хотя жир обеспечивает большую энергию и, следовательно, способствует ожирению, обеспечивая избыточные калории, сахар чаще ассоциируется с модуляцией привычного переедания и, следовательно, с поведением, подобным зависимости, связанным с ожирением (Shearrer et al., 2018a).
Чрезмерное потребление в подростковом возрасте приводит к долгосрочным изменениям в системе дофаминергического вознаграждения и может вызывать сенсибилизацию, вызванную стимулами, которая наблюдается во взрослом возрасте (Naneix et al., 2018; Robinson and Berridge, 2008).
Сенсибилизация, вызванная стимулами, относится к усилению эффекта стимулов после повторных воздействий, например, повторное воздействие громкого шума может вызвать сенсибилизацию к шуму, вызывая усиленную реакцию, то же самое относится к повторным воздействиям наркотика, вызывающего злоупотребление.
При наркомании сенсибилизация вызывает изменения в передаче дофамина и экспрессии дельта-FosB, что способствует усилению тяги и рецидиву (Robinson and Berridge, 2001).
Крысы-подростки, которым разрешалось потреблять сахарозу (5% раствор сахарозы) в течение шестнадцати дней, позже были протестированы на мотивацию к поиску сахарина, мальтодекстрина или кокаина (Vendruscolo et al., 2010).
Взрослые крысы показали снижение мотивации к приобретению сахарина и мальтодекстрина, однако это не относится к кокаину (Vendruscolo et al., 2010). В дополнение к этим результатам, крысы, которым был предоставлен выбор между раствором сахарина и внутривенным кокаином, продемонстрировали, что вознаграждение, предоставляемое сладостью, превзошло желание кокаина (Lenoir et al., 2007); продолжаются жаркие дебаты о том, вызывает ли сахар привыкание на самом деле (Benton, 2010; Corsica and Pelchat, 2010; Wilson, 2010).
Была выдвинута гипотеза, что аддиктивное поведение, присутствующее после длительного потребления сахарозы, возникает из-за очень вкусной природы сахара, а не из-за нейрохимических эффектов самого сахара (для обзоров см. (Ahmed et al., 2013; Hoebel et al., 2009; Westwater et al., 2016; Avena et al., 2009; Criscitelli and Avena, 2017; Kendig, 2014)).
Целью этого обзора является не обсуждение аргумента, а изучение роли сахара в эмоциональной дисфункции.
Общая нейрохимия лежит в основе потребительского поведения и эмоций
Исследования нейровизуализации и модели животных, изучающие механизмы, вовлеченные в прогрессирование ожирения, выявили нейробиологические корреляты, участвующие в нейроадаптации ожирения и потребления сахара (Lindgren et al., 2018; Novelle and Diéguez, 2018).
Доклинические модели потребления на грызунах полезны для изучения нейронных областей и путей, лежащих в основе потребительского поведения (Holgate et al., 2017; Bonin et al., 2018)
при этом показано, что прерывистый доступ к сахарозе оказывает влияние на опиоидные, холинергические и, что важно, дофаминергические рецепторы (Shariff et al., 2016; Avena et al., 2008; Colantuoni et al., 2002; Pratt and Kelley, 2004).
Опиоиды
Опиоидные рецепторы экспрессируются по всей лимбической системе и играют ключевую роль в регуляции страха, счастья, гнева, возбуждения, мотивации и кормления, связанного с вознаграждением (Nummenmaa and Tuominen, 2017; Levine and Billington, 2004).
Поведение, вызванное опиоидами, включает модуляцию боли, наркотической зависимости и контроль автономной нервной системы, которая включает эмоции, когнитивные процессы и преодоление стресса (Benarroch, 2012; Belzeaux et al., 2018).
Опиоиды влияют на то, как мы обрабатываем вознаграждения, посредством взаимодействия с дофаминовой системой (Bray and Bouchard, 2014).
о, как животное ведет себя по отношению к еде, также модулируется эндогенными опиоидными нейротрансмиттерами, которые являются нейронными субстратами, создающими восприимчивость к вознаграждению (Wise, 1996; Kavaliers and Hirst, 1987).
Связь между стрессом, уровнем кортизола и потреблением сахара далека от понимания, однако, по-видимому, диеты с высоким содержанием сахара играют важную роль в регуляции оси HPA и переедании, связанном со стрессом.
Совсем недавно наша лаборатория выяснила влияние варениклина (одобренного FDA препарата для снижения тяги к никотину) на потребление сахарозы у крыс (Shariff et al., 2016).
Варениклин, частичный агонист подтипов α4β2* и антагонист α6β2* nAChRs, значительно снизил потребление сахарозы как при краткосрочном, так и при долгосрочном запое (прерывистый доступ) употреблении алкоголя. В то время как участки связывания α4β2* nAChR были увеличены, α6β2* nAChR были значительно уменьшены в NAc в результате как краткосрочного, так и долгосрочного потребления сахарозы (Shariff et al., 2016).
Путем модуляции высвобождения дофамина в NAc (Feduccia et al., 2014) контроль потребления сахарозы никотиновыми рецепторами и их подтипами может обеспечить многообещающие терапевтические стратегии при ожирении (Cocores and Gold, 2008).