В обычных учебниках, да и в любых книгах по растительному разнообразию, характеристика таксона до сих пор начинается с морфологии. Это было вполне естественным на морфологическом этапе систематики. Однако современная систематика построена, в первую очередь, на структуре генома. Вся остальная информация рассматривается лишь как дополнительно-иллюстративная. Давайте и мы попробуем начать с генома. В прошлом году мы уже чуток касались этого вопроса, когда обсуждали голосеменные в целом, и установили следующее.
Геном голосеменных, если сравнивать его с геномом цветковых, огромный, неповоротливый, не динамичный. В нем много лишнего и бесполезного: разного рода генетического мусора. Образно это называется «genom obesity», ожирение генома. Большую часть «жира» составляют мобильные генетические элементы – небольшие фрагменты ДНК, которые способны самопроизвольно вырезаться (копироваться) в одном месте и вставляться в другом. Хвойные по структуре генома являются даже не типичными, а наиболее яркими представителями голосеменных. Из-за экономической важности их геном неплохо изучен. На их примере мы рассмотрим кое-какие подробности и углубим тем самым наши молекулярно-биологические знания.
Начнем с числа хромосом в гаплоидном наборе (1n). У хвойных диапазон разнообразия совсем маленький: от 9 до 19. При этом крайние варианты встречаются редко: большинство видов имеют 11 (Cupressaceae), 12 (Pinaceae) или 13 (Araucariaceae) хромосом. Малое разнообразие означает крайнюю редкость хромосомных перестроек, консервативность геномной организации. Это характеризует стабильность хвойного генома и резко контрастирует с ситуацией у цветковых, где разнообразие многократно больше: и вообще, и внутри семейств, и даже внутри родов.
Хромосомы у хвойных довольно крупные. Их неплохо видно даже в простой световой микроскоп. Вслед за подсчетом их числа уже в начале прошлого века исследователи начали их нумеровать, измерять, зарисовывать (фотографировать). Оказалось, что идентифицировать каждую, особенно у разных видов, не так-то просто: многие очень похожи, как сосиски в супермаркете. Проблема решилась с изобретением флуоресцентной гибридизации in situ (fluorescent in situ hybridization = FISH) в 1993 г. В технические подробности мы, естественно, вникать не будем. А суть ее в том, что специально разработанными светящимися метками помечаются идентичные по последовательности нуклеотидов участки ДНК. Хромосомы от этого становятся яркими, пестрыми, красивыми, а главное – легко отличимыми. Ниже приведена картинка из вот этой работы: Shibata F., Matsusaki Y. & Hizume M. (2016). A comparative analysis of multi- probe fluorescence in situ hybridization (FISH) karyotypes in 26 Pinus species (Pinaceae). Cytologia. Vol. 81. N4. P. 409–421.
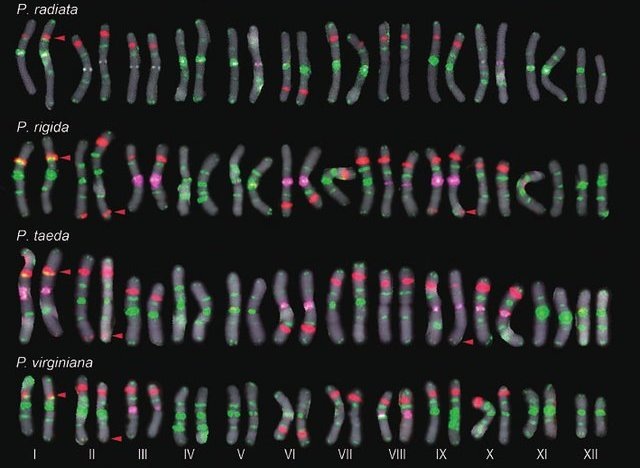
Как видите, все пары хромосом каждого вида расставлены строго по порядку с 1-й по 12-ю. И виды, и пары безошибочно распознаются этим методом. Бери и изучай каждую хромосому в отдельности!
Полиплоидия чрезвычайно характерна для соседей хвойных по эволюционной лестнице: соседа снизу - папоротников (рекорд - Ophioglossum reticulatum 96n, 1440 хромосом) и соседа сверху - цветковых (рекорд - Sedum suaveolens, 80n, 640 хромосом). У хвойных нет ничего похожего. Абсолютное большинство видов – обыкновенные диплоиды. В их вегетативных клетках двойной набор хромосом (2n): один комплект от папы, другой от мамы. До самого последнего времени было известно лишь два исключения, оба в семействе Cupressaceae: тетраплоидная (4n) Fitzroya из Южной Америки и гексаплоидная (6n) Sequoia из Америки Северной.
Однако в 2019 г. вышла вот такая публикация: Farhat P, Hidalgo O, Robert T, Siljak-Yakovlev S, Leitch I, Adams RP, Bou Dagher Kharrat M. Polyploidy in the conifer genus Juniperus: an unexpectedly high rate. Front Plant Sci. 2019; 10: 676. В ней показано, что редкость полиплоидии у хвойных несколько преувеличена. В роде можжевельник (Juniperus) при ближайшем рассмотрении 16 видов оказались тетраплоидами (4n), а один даже и гексаплоидом (6n). Это можжевельник вонючейший (J. foetidissima), он есть в Крыму и на Черноморском побережье Кавказа. Полиплоидия широко распространена у процветающих цветковых. Возможно, она также способствовала и эволюции Juniperus – одного из наиболее успешных родов хвойных.
Собственно геном – это вся ДНК данного вида. На рубеже настоящего и прошлого веков в науке, изучающей наследственность, произошла революция: впервые были полностью секвенированы геномы нескольких видов. Слово секвенирование происходит от английского sequence – последовательность. Секвенировать, значит определять последовательность нуклеотидов ДНК. В не научных СМИ обычно употребляют глагол «расшифровать». Первыми организмами с «расшифрованным» геномом были человек и арабидопсис (небольшое травянистое растение с малюсеньким геномом, 125 миллионов пар нуклеотидов (млн.п.н.), которое впоследствии стало модельным для исследования растительных геномов). С этого момента генетика, которая изучала отдельные гены, постепенно трансформировалась в ГЕНОМИКУ. Из деревьев первым в 2006 г. секвенировали геном американского вида Populus trichocarpa (тополя волосистоплодного), очень маленький, чуть менее 500 млн.п.н. С этого времени тополь стал «древесным арабидопсисом» - модельным родом для исследования генома деревьев.
О гигантском размере генома хвойных мы уже вспоминали. Действительно, это самые крупные растительные геномы. Технические возможности для их секвенирования появились примерно к 2010 году. Началась гонка трех проектов: шведского (Picea abies, ель европейская), канадского (Picea glauca, ель канадская) и американского сразу по трем видам (Pinus taeda, сосна ладанная; Pinus lambertiana, сосна Ламбера; Pseudotsuga menziesii, псевдотсуга мингиса). Как это ни странно, победили шведы в 2013 г. Правда, как потом выяснилось, они слегка смухлевали: секвенировали лишь 60% генома. Однако всё равно остались в истории. Геномы остальных 4 видов были сделаны на более высоком уровне. Что интересного удалось обнаружить. Об этом мы узнаем в следующий раз.