Размер генома – предмет сложный. С одной стороны, понятно, что у простых организмов, например, у водорослей или бактерий, он в среднем многократно меньше (миллионы пар нуклеотидов), чем у сложных, например, у цветковых или млекопитающих (миллиарды пар нуклеотидов = млрд.п.н.). С другой стороны, в слишком большом геноме тоже нет ничего хорошего. Ведь при каждом клеточном делении его надо весь (весь!!!) удваивать. Большой геном – это большие затраты вещества и энергии. Всё хорошо в меру. Размер генома у разных групп наземных растений сравнили вот в этой научной публикации: Ilia J. Leitch and Andrew R. Leitch. Genome size diversity and evolution in land plants // I.J. Leitch et al. (eds.), Plant Genome Diversity Volume 2, DOI 10.1007/978-3-7091-1160-4_19, # Springer-Verlag Wien 2013. P. 307-322:
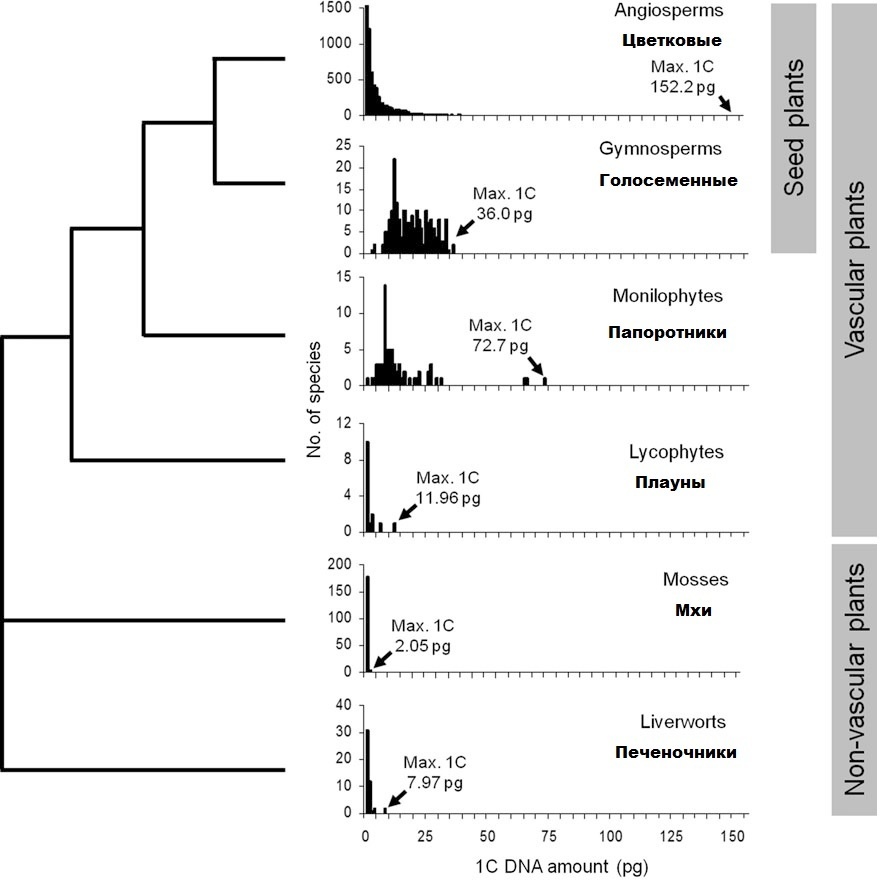
Слева филогенетическое древо высших растений: 6 основных подклассов. Справа – гистограммы: по горизонтали размер генома, только не в парах нуклеотидов, а в пикограммах (1 пг это примерно 1 млрд. нуклеотидов); по вертикали – число видов с известным науке размером генома. Как видите, у бессосудистых подклассов (печеночники, мхи) геном небольшой, у высших споровых (плауны, папоротники) – значительно больше, у голосеменных – максимальный, у цветковых – ОПТИМАЛЬНЫЙ. Приведем некоторые подробности:
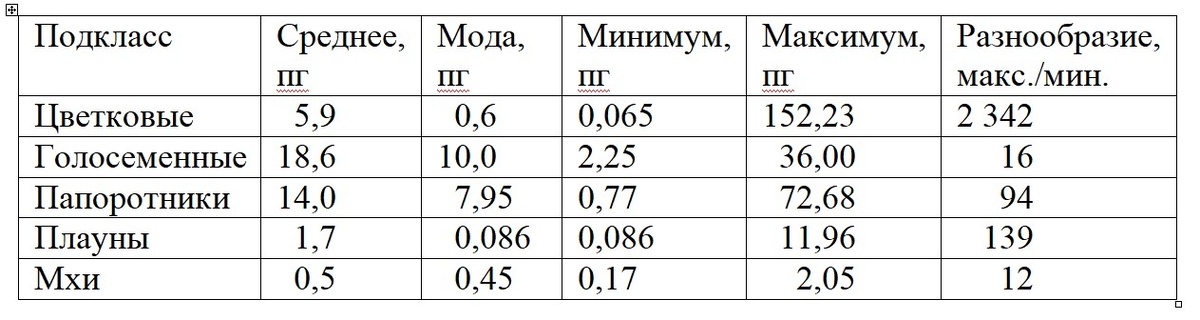
В таблице хорошо видна криволинейная связь между эволюционной «продвинутостью» и размером генома: от мхов к голосеменным он сильно увеличивается, а от голосеменных к цветковым резко снижается. Обратите внимание на моду. Так называют значение признака, которое чаще всего встречается в вариационном ряду. У голосеменных самый «модный» размер генома – 10 пг, у цветковых – почти в 20 раз меньше. Абсолютное большинство последних имеют маленький геном и лишь немногие - огромный. У голосеменных, наоборот, преобладают средние значения признака. Вызывают интерес данные о разнообразии размеров генома внутри подклассов. Оно минимальное у мхов, резко возрастает к плаунам, потом снова снижается к голосеменным до очень низкого уровня и, наконец, возрастает до запредельной величины у цветковых. Последним принадлежат оба рекорда: и самый маленький, и самый большой геном среди наземных растений.
Получается, закономерность всё-таки есть, но сугубо в среднем. У конкретных видов, родов, семейств всё может быть с точностью до наоборот. Достаточно привести пример из животного мира: у одного из видов амебы (Amoeba dubia) размер генома 680 000 млрд.п.н., у человека (Homo sapiens) – 3 млрд.п.н., т.е. в 227 раз меньше. Вот вам и венец творения! Таких исключений полным-полно и в мире растений: см. приведенную выше таблицу.
Как объяснить, интерпретировать всё это? Низшие растения (водоросли) имеют очень маленький геном. Этого вполне достаточно для их незамысловатой жизни в благоприятной водной среде. Выход растений на сушу потребовал резкого усложнения структурно-функциональной организации тела, которое было немыслимо без существенного увеличения размеров генома. Появились и начали проходить испытание разные механизмы увеличения: полиплоидизация, дупликация (удвоение) больших и малых фрагментов ДНК, активизация мобильных генетических элементов. Система разгонялась медленно. Поэтому у мхов геном еще небольшой и не особо изменчивый. Всё это происходило методом проб и ошибок. Были удачные опыты, были неудачные. Поэтому у плаунов и папоротников наблюдается большое разнообразие по размеру генома. На полную мощность процесс вышел у голосеменных: у них огромные, но минимально разнообразные по размеру геномы.
Пока увеличение количества ДНК шло на пользу эволюции, она всерьез не «заботилась» о разработке механизмов сокращения (оптимизации) размеров генома. Естественный отбор устранял лишь вредные мутации, безвредные не трогал. Последние, накапливаясь в большом количестве, превратились в проблему. Ко времени появления цветковых она встала во весь рост. Встала и была решена. Геномы цветковых стали гибкими, динамичными, эффективными. В них быстро и легко происходили необходимые изменения, в том числе, через увеличение количества ДНК, но они гармонично дополнялись механизмами самоорганизации, очистки от балласта, мусора, отходов геномной жизнедеятельности. Геном в таком совершенном состоянии показал чудеса эволюции, создал поразительное разнообразие форм, а также и разнообразие самих геномов, в том числе, по размеру.
Размер генома, конечно, влияет на эволюцию не сам по себе. Однако факты – упрямая вещь. На примере цветковых установлено, что виды с большим геномом почти всегда имеют ограниченный диапазон фенотипического разнообразия, жизненных стратегий и экологических вариантов. Возьмем самую простую вещь: минеральное питание. ДНК состоит из нуклеотидов. Нуклеотид – это азотистое основание + углевод дезоксирибоза + фосфатная группа. При каждом клеточном делении количество ДНК удваивается. Поэтому чем больше геном, тем больше потребности в азотном, а особенно в фосфорном питании. Понятно, что при прочих равных условиях у растений с большим геномом ниже конкурентоспособность, следовательно, и эволюционный потенциал.
Это с одной стороны. С другой стороны, размер размеру рознь. У покрытосеменных геномы были сформированы преимущественно несколькими событиями дупликации целого генома (полиплоидизации). У хвойных таких «событий» не было. Был медленный, но неуклонный процесс: размер генома увеличивался в течение миллионов лет в результате массивного накопления относительно единообразных мобильных генетических элементов (МГЭ). Приведем такую аналогию. Вот есть у нас один хороший работник. Его вес 75 кг. Пришел еще один точно такой же. Их общий вес 150 кг. Они сделали работы в 2 раза больше. Теперь другая ситуация: наш 75-килограммовый работник растолстел до 150 кило. Понятно, что ему уже не до работы. Так вот, первый случай – это условно полиплоидия, характерная для цветковых, второй – «геномное ожирение», о котором мы говорили выше применительно к хвойным. Аналогия, конечно, не полная. Зато понятная. Посмотрим картинку из вот этой научной публикации: Yang Liu and Yousry A. El-Kassaby. Novel insights into plant genome evolution and adaptation as revealed through transposable elements and non-coding RNAs in conifers. Genes 2019, 10, 228; doi:10.3390/genes10030228.
На верхней диаграмме показан размер генома. Для хвойных данные по 4, для лиственных по 38 видам. Различия, как видите, 20-кратные. На нижней диаграмме число генов (белок-кодирующих транскриптов): различий между хвойными и лиственными почти нет. Это означает, что у хвойных на один реально работающий ген приходится примерно в 20 раз больше всякого генетического хлама, чем у лиственных. Кодирующие белок последовательности ДНК у хвойных составляют всего лишь 0,1% генома! Это очень мало.
Какова сравнительная эффективность двух типов геномов в части адаптации и эволюции? Если понимать эти процессы по-простому, по-старому, по-дарвиновски, то они основаны на естественном отборе. Чем больше разнообразие, тем больше возможностей для отбора. Разнообразие на генетическом уровне создается мутациями и рекомбинацией. Про мутации мы уже говорили, когда обсуждали голосеменные в целом. У голосеменных, в том числе, и у хвойных, скорость молекулярной эволюции (число мутаций за единицу времени) примерно в 7 раз ниже, чем у цветковых. Теперь несколько слов о рекомбинациях.
Рекомбинация - это процесс, который обеспечивает перемешивание генов в ряду поколений. При формировании половых клеток гены, полученные от родителей, “перетасовываются”: две гомологичные хромосомы обмениваются фрагментами. При оплодотворении гены двух родителей случайно комбинируются в зиготе. Сочетание этих двух случайных процессов, наряду с мутациями, обеспечивает уникальность набора генов каждого организма, создает разнообразие – пищу для естественного отбора. Рекомбинация как обязательный процесс, предшествующий образованию половых клеток, возникла давно. Поэтому почти у всех современных живых организмов, кроме бактерий, она происходит примерно одинаково. Одинаково, но с разной скоростью. Хвойные и тут сильно отстают от цветковых. Информация взята вот из этой научной публикации: Juan P Jaramillo-Correa, Miguel Verdú, Santiago C González-Martínez. The contribution of recombination to heterozygosity differs among plant evolutionary lineages and life-forms // Evolutionary Biology. 2010. V. 10 (22).
Черными кругами показана скорость рекомбинации: больше круг – больше скорость. Цветковые довольно разнообразны в этом отношении, но круг есть почти у всех. У большинства хвойных круга нет, есть только едва заметная точка. Такого низкого уровня рекомбинации нет больше ни у каких многоклеточных организмов. Он низок не только вообще, в расчете на целый геном, но и в расчете на совокупность реально функционирующих генов. Это сильно снижает скорость эволюции: и напрямую, и через увеличение размера генома (потому, что рекомбинация – главный способ утилизации «генетического мусора»).
Обычно считается, что голосеменные в эволюционном смысле старше цветковых чуть не на 200 миллионов лет. Так да не так. Ведь если это последняя развилка древа наземных растений, то возраст обеих ветвей должен быть одинаковым. Другое дело, что все эти 200 млн. лет цветковые не были цветковыми. Они были одной из групп семенных папоротников, которые «готовились» стать цветковыми. Но ведь сути дела-то это не меняет. Эти семенные папоротники явно не спешили, «играли вдолгую», исподволь накапливали потенциал, в первую очередь, отлаживали разнообразные механизмы эффективной трансформации генома. Потом «выстрелили»: за очень короткий по геологическим меркам срок дали гигантское разнообразие форм. Приведем лишь один, пожалуй, самый распространенный и самый эффективный способ видообразования у цветковых (Yang Liu and Yousry A. El-Kassaby. Novel insights into plant genome evolution and adaptation as revealed through transposable elements and non-coding RNAs in conifers. Genes 2019, 10, 228; doi:10.3390/genes10030228):
Как понять эту схему? Предковый диплоидный вид породил два новых диплоидных вида (2n). Не важно каким способом: у цветковых их много. Между этими двумя видами произошла гибридизация: возникли тетраплоидные гибриды (4n). Они сразу же оказались полностью изолированы от родительских видов из-за разной плоидности.
Гибридизация + полиплоидизация – это стресс для генома: гибриды как бы угодили в черный ящик с бурными генетическими процессами. Стресс активизировал мутации и рекомбинации. Мутации вызывали нарастание различий между гомологичными хромосомами. Они стали активно рекомбинировать: обмениваться генетическим материалом. При этом формировались новые или модифицированные гены, а также новые генные комплексы. Всё полезное по возможности поддерживалось естественным отбором, всё вредное и лишнее элиминировалось из генома.
Гомологичные хромосомы с каждым новым поколением становились все меньше похожи друг на друга. Это значит, что был запущен процесс диплоидизации, т.е. движение к такому состоянию, когда число хромосом стало в 2 раза больше, чем у предкового вида, но они были все разные по структуре и содержанию (на рисунке это показано разными цветами: см. три кружка на правом краю). Особи при этом группировались в популяции по географическому и экологическому признакам, а главное – по признаку репродуктивной изоляции, которая у цветковых образуется довольно быстро, притом, не столько на экологическом, сколько на генетическом уровне. Каждая такая популяция стала зародышем нового вида.
Красиво? Красиво! Так вот: никакой такой красоты у хвойных нет и быть не может. Потому, что полиплоидия у них не развита, все виды мутаций и рекомбинаций происходят на порядок медленнее, репродуктивная изоляция возникает еще медленнее. Принцип их эволюции был совсем другой. Он не требовал длительной подготовки, оттачивания тонких и точных генетических механизмов, но при этом давал неплохой результат. Хвойные пошли по пути быстрого успеха.
Что же это был за принцип? В качестве главного инструмента эволюции они использовали МГЭ! Да, те самые, которые мы чуть выше обозначили как мусор и балласт. На самом деле, это не совеем верно. Они даже и сейчас нередко работают на пользу «хозяевам». А уж в те далекие мезозойские времена, когда геном хвойных был молодым и «поджарым», не отягощенным «жировыми отложениями», они буквально рулили процессом. Чтобы разобраться с этим парадоксом, нам придется поближе познакомиться с МГЭ. Кому это не интересно, того мы к чтению не принуждаем. Этот раздел можно спокойно пропустить. Для остальных продолжим. В следующий раз.