
Выкладываем перевод статьи опубликованной в журнале Nutrients в 2017 году в которой ученые рассказывают о серии научных экспериментов которой они ответили на вопрос в чем будет отличие переваривания организмом человека выпечки из однозернянки от выпечки из обычной мягкой пшеницы. С помощью аппарата имитирующего работу нашего ЖКТ, ученые "переварили" продукт и зафиксировали чем конкретно выпечка из однозернянки может быть полезна для нашего здоровья. Перевод с английского. Ссылка на оригинал: https://www.mdpi.com/2072-6643/9/11/1232
Комплексная оценка пользы для здоровья хлебобулочных изделий на основе однозернянки
Аннотация: в настоящее время признана высокая питательная ценность цельного зерна, и растет интерес к древним сортам для производства цельнозерновых пищевых продуктов с улучшенными питательными свойствами. Среди древних зерновых культур хорошей альтернативой может стать однозернянка. В этой работе мука из однозернянки была проанализирована на содержание в ней каротиноидов, свободных и связанных фенольных кислот и проведено сравнение с пшеничной мукой. Наиболее многообещающие виды муки были использованы для производства обычного хлеба и хлеба на закваске. Хлеб был подвергнут перевариванию в лабораторных условиях и охарактеризован до и после переваривания. Были отобраны четыре вида хлеба, обладающие наилучшими характеристиками, и продукт их переработки был использован для оценки их противовоспалительного действия с использованием клеток Caco-2. Наши результаты подтверждают более высокий уровень каротиноидов в однозернянке, чем в современной пшенице, и эффективность ферментации закваски для поддержания этого уровня, несмотря на более длительное воздействие атмосферного кислорода. Более того, в культивируемых клетках хлеба из однозернянки был обнаружен противовоспалительный эффект, хотя и замаскированный воздействием пищеварительной жидкости. Это исследование представляет собой первую комплексную оценку потенциальной пользы для здоровья хлебобулочных изделий на основе однозернянки по сравнению с хлебобулочными изделиями на основе мягкой пшеницы и расширяет наши знания о древних злаках.
Ключевые слова: однозернянка; пшеница; каротиноиды; хлеб; фенольные кислоты; ферментация закваски; переваривание in vitro; клетки Caco-2; противовоспалительный эффект.
1. Введение
Несколько исследований показали четкую корреляцию между потреблением цельнозерновых продуктов и снижением риска сердечно-сосудистых заболеваний [1,2], диабета [3] и некоторых видов рака [4]. Полезные свойства цельнозерновых продуктов в основном объясняются их содержанием микроэлементов и фитохимических веществ [5-7]. Злаки являются одними из самых богатых продуктов питания фенольными кислотами, их содержание сравнимо или даже выше, чем в ягодах, фруктах и овощах [8]. Кроме того, некоторые злаки богаты лютеином и зеаксантином [9,10]. Микроэлементы и фитохимические вещества в основном сосредоточены во внешних слоях зерен [11], и это может объяснить профилактический эффект, связанный с высоким потреблением цельнозерновых продуктов [12].
В настоящее время признана более высокая питательная ценность цельнозерновой муки по сравнению с рафинированной [13], и растет интерес к древним культурам как источнику цельнозерновой муки [14].
Однозернянка (Triticum monococcum L. ssp. monococcum) - древняя сельскохозяйственная культура. По сравнению с полиплоидной пшеницей, она отличается более высоким содержанием белков, полиненасыщенных жирных кислот, фруктанов и фитохимических веществ, таких как токолы, каротиноиды, алкилрезорцины, фитостеролы, и более низкой активностью α-, β-амилазов и липоксигеназов [15]. Кроме того, в однозернянке содержится очень мало глютеновых пептидов, стимулирующих Т-клетки [16]. Однозернянка может стать хорошей альтернативой для производства функциональных хлебобулочных изделий.
В хлебобулочных изделиях обработка может способствовать повышению функциональности [17,18]. Было показано, что ферментация закваски, включающая взаимосвязь между метаболизмом микроорганизмов и ферментами злаковых, значительно влияет на функциональные свойства хлебобулочных изделий на закваске [19]. Этот тип ферментации может привести к образованию новых питательных молекул, таких как функциональные пептиды и производные аминокислот [20,21], которые образуются либо в результате гидролитической активности бактерий [20], либо в результате их собственного синтеза [22]. Чтобы оказывать положительное воздействие на организм человека, биологически активные соединения должны гидролизоваться из пищевой матрицы и всасываться в кишечнике. Биодоступность биологически активных соединений, т.е. процентное содержание, выделяемое из пищевой матрицы и доступное для усвоения слизистой оболочкой кишечника, является важным параметром, на который может влиять множество различных факторов, включая пищевую матрицу и процесс обработки пищевых продуктов [23,24]. Ферментация молочнокислыми бактериями может улучшить биодоступность питательных веществ и привести к образованию соединений, обладающих антиоксидантной и противовоспалительной активностью [19]. Сообщалось, что молочнокислые бактерии, содержащиеся в закваске, выделяют или синтезируют антиоксидантные и противовоспалительные пептиды во время ферментации муки из злаков [20].
В этой работе были проанализированы различные виды пшеничной муки и муки из однозернянки на предмет содержания в них каротиноидов и фенольных кислот. Были отобраны самые богатые этими функциональными соединениями сорта муки, которые использовались для выпечки хлеба с использованием двух различных методов ферментации (обычной и на закваске).
Хлеба были переработаны in vitro (в пробирке) с использованием динамического желудочно-кишечного дигестатора и охарактеризованы до и после переваривания. На основе обобщенных результатов были отобраны четыре вида хлеба, и продукт их переваривания в кишечнике был добавлен в кишечные клетки Caco-2. Клетки подвергались воспалительному стрессу, и оценивалось влияние добавок на различные маркеры воспаления.
В целом, в этом исследовании оценивалось, как тип муки и способ ферментации могут влиять на питательные свойства хлеба, а также на биодоступность и противовоспалительное действие его функциональных соединений. Сочетание различных результатов дает комплексное представление о возможной пользе хлеба на основе однозернянки для здоровья.
2. Материалы и методы
2.1. Материалы
Фенольные кислоты (4-гидроксибензойная кислота, кофейная кислота, хлорогеновая кислота, феруловая кислота, галловая кислота, p-Кумаровая кислота, синаповая кислота, сирингиновая кислота и транс-коричная кислота) стандарт чистоты (чистота ≥99,5%) в виде порошка; метанол, ацетонитрил, ацетон, диэтиловый эфир, пригодный для ВЭЖХ (Высокоэффективной жидкостной хроматографии), этилацетат и вода; фосфорная кислота (85-87% по массе), соляная кислота (37% по массе), одноосновный фосфат натрия (≥98%), гранулы гидроксида натрия (≥98%) были произведены компанией Sigma-Aldrich Italia(Милан, Италия). Исходные растворы фенольных кислот (1 мг/мл) готовили путем растворения 10 мг каждого чистого вещества в 10 мл метанола. Стандартные растворы получали путем разбавления исходных растворов подвижной фазой (исходный состав). Сертифицированные исходные растворы каротиноидов в ацетонитриле (520 мкг/мл неоксантина; 752 мкг/мл виолаксантина; 590 мкг/мл антераксантина; 830 мкг/мл зеаксантина; 771 мкг/мл лютеина; 1033 мкг/мл β-криптоксантина; 894 мкг/мл β-каротина) были приобретены в DHI (Херсхольм, Дания) и разбавлены метанолом перед инъекцией. При хранении при температуре -20 °C в темноте исходные растворы были стабильны в течение как минимум одного месяца (по оценке ВЭЖХ); стандартные растворы готовили свежими каждый день.
Были использованы XS Instruments pH 50 pHmeter (Капри, Италия), центрифуга Thermo Scientific CL10, ножевая мельница IKA A11 Basic (Штауфен, Германия) и сублимационная сушилка M. Christ Alpha 1–4 LD.
Клетки Caco-2 и HEK-Blue™ IL-6 были получены от Sigma-Aldrich (Сент-Луис, Миссури, США) и InivoGen (Сан-Диего, Калифорния, США), соответственно. Сбалансированный солевой раствор Эрла (EBSS), модифицированная среда Игла Дульбекко (DMEM), фетальная бычья сыворотка (FBS), пенициллин, стрептомицин, L-глютамин были получены от GE Healthcare (Литтл-Чалфонт, Великобритания). Незаменимые аминокислоты (NEAA), пируват натрия и фунгизон были приобретены в компании Thermo Fisher Scientific (Уолтем, Массачусетс, США).
2.2. Образцы муки
Очищенная пшеничная мука (Triticum aestivum L.; стандартная мука, SF), в том числе двух испанских сортов (550 S and 650 S) и двух польских сортов (550 P and 650 P), была предоставлена компаниями INDESPAN (Валенсия, Испания) и VINI (Рогозник, Польша) соответственно. Мука из однозернянки (или древняя мука, AF), включая органическую цельнозерновую и рафинированную муку, полученную из Triticum monococcum L. var. Monlis, была предоставлена компанией Prometeo (Урбино, Италия).
2.3. Штаммы молочнокислых бактерий (Lactic Acid Bacteria (LAB)) и приготовление закваски-стартера
Были использованы штаммы Lactobacillus plantarum 98a, Lactobacillus sanfranciscensis BB12, Lactobacillus brevis 3BHI, принадлежащие департаменту сельскохозяйственной и пищевой науки и технологии (DISTAL) Болонского университета. Лабораторные штаммы выращивали отдельно в бульоне Man Rogosa Sharpe (MRS) (Oxoid, Милан, Италия) при температуре 37 °C в течение 24 ч. Клетки собирали центрифугированием при 4000×g в течение 10 мин и дважды промывали стерильной водой. Для приготовления закваски 600 г муки осторожно смешивали с 270 мл воды и вносили по 80 мл водной суспензии каждого отдельно выращенного штамма. Замешенное тесто выдерживали при температуре 30 °C в течение 24 ч. для получения зрелой закваски.
2.4. Процессы ферментации и выпечки
Были приняты 2 типа ферментационных процессов: (i) обычное брожение (CF) на основе коммерческих прессованных дрожжей (хлебопекарных дрожжей), (ii) брожение на закваске, полученной из пшеничной муки (SSF) или муки однозернянки (SAF). Традиционную закваску проводили с использованием хлебопекарных дрожжей (2,5% муки) при температуре 30 °C в течение 1,5 ч. Закваску для брожения теста получали путем добавления описанной выше закваски (около 30% от общего количества теста). Содержание инокулята в конечном тесте составляло приблизительно 4 × 108 CFU/g (КОЕ/г) для каждого штамма молочнокислых бактерий (LAB). В готовое тесто добавляли хлебопекарные дрожжи и сбраживали, как описано для обычного брожения. Добавляли воду до получения DY = 250 и соль в количестве 2 г на 100 г муки. Ферментированное тесто выпекали при температуре 195 °C в течение 45 минут в промышленной печи фирмы VINI (Рогозник, Польша).
Таким образом, было получено шесть экспериментальных сортов хлеба (таблица 1).
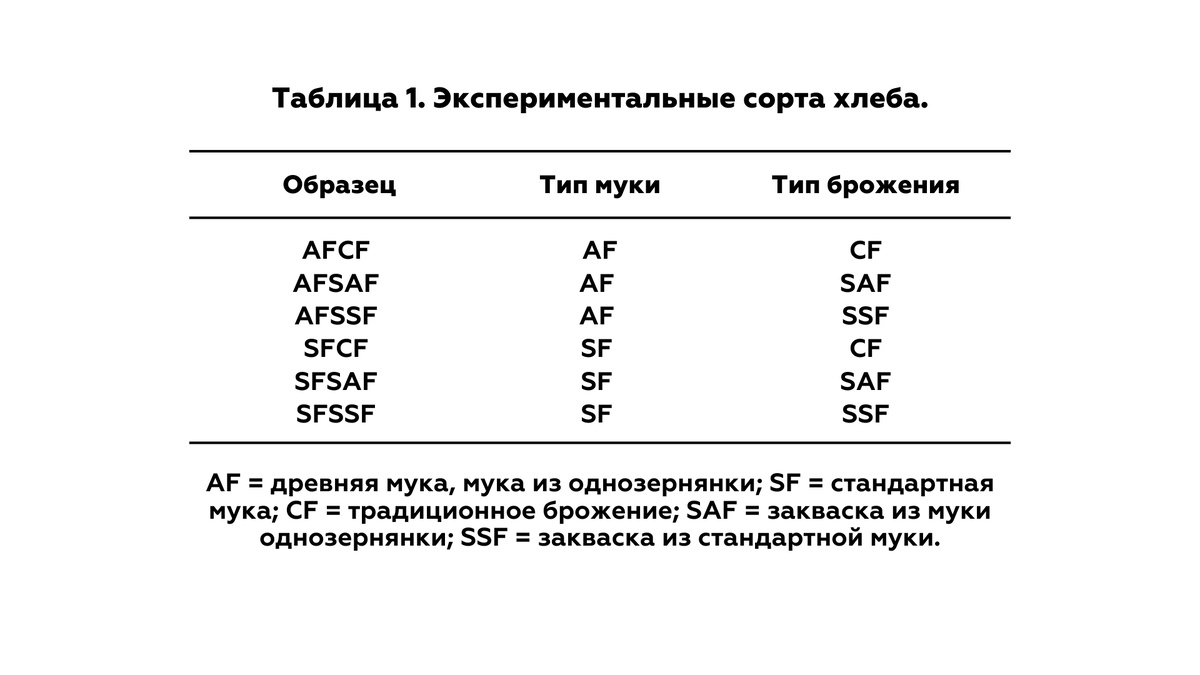
2.5. Переработка образцов хлеба в лабораторных условиях
Шесть экспериментальных сортов хлеба были переработаны in vitro с использованием динамического желудочно-кишечного дигестатора (DGD). DGD - это многокомпонентная система с компьютерным управлением, которая имитирует биологическую среду в желудке и тонком кишечнике человека [25]. Процесс переваривания проводили на 50 г экспериментального хлеба или 50 г воды (холостое переваривание) в течение 360 мин (120 мин в желудке и 240 мин в тонком кишечнике) при температуре 37 °C. Она включала в себя несколько последовательных ферментативных процедур: выделение слюны (α-амилаза), желудочной секреции (пепсин и липаза) при кислом рН и выделение кишечника (желчь и панкреатин) при нейтральном рН. Секреция пищеварительных соков, регулирование рН и постепенное опорожнение желудка и тонкого кишечника были смоделированы в соответствии с физиологическими данными [26,27]. Чтобы свести к минимуму фотоокисление, все эксперименты проводились при постоянной промывке газообразным азотом и в защищенном от света месте. Полученный при каждом переваривании кишечный дигестат центрифугировали при 5000× g при 4 °C в течение 45 мин, а надосадочные жидкости фильтровали через целлюлозные фильтры с порами 0,22 мкм для получения водных фракций, содержащих биодоступные аналиты [28]. Дигесты замораживали и хранили при температуре -20 °C до их анализа.
2.6. Определение содержания фенольной кислоты методом ВЭЖХ
Образцы хлеба подвергали сублимационной сушке и тонкому измельчению в ножевой мельнице в течение 4 периодов по 30 секунд. Образцы муки непосредственно использовали для экстракции, в то время как дигесту in vitro перед экстракцией подвергали сублимационной сушке. Фенольные кислоты экстрагировали из образцов в соответствии с протоколом, разработанным Moore и соавт. [29], с изменениями. 2-граммовую аликвоту порошкообразного образца помещали в ножевую мельницу с 20 мл смеси метанол/ацетон/вода (7/7/6, v/v/v); после 30-секундного перемешивания смесь центрифугировали в течение 5 мин при 1400×g. Были выделены три фракции: две (свободная и растворимая конъюгированная) из надосадочной жидкости образца и одна (нерастворимая связанная) из осадка. В обработанных образцах осадка не было, поэтому из них были выделены только две фракции (свободная и растворимая конъюгированная).
2.6.1. Свободные фенольные кислоты (СВОБОДНАЯ фракция)
Один мл надосадочной жидкости смешивали с 10 мл of 0.01 N HCl и трижды экстрагировали равным объемом смесь диэтилового эфира/этилацетата (1/1, v/v). Органические экстракты сливали, высушивали во вращающемся испарителе и повторно растворяли в 2 мл метанола.
2.6.2. Растворимые конъюгированные фенольные кислоты (фракция SC)
Восемь мл надосадочной жидкости смешивали с 2 мл 10 M NaOH и гидролизовали в потоке азота при постоянном перемешивании в течение 1 часа. Затем раствор подкисляли до pH 2 с 12 M HCl и подвергали процедуре экстракции, уже описанной для СВОБОДНОЙ фракции.
2.6.3. Нерастворимые связанные фенольные кислоты (фракция IB)
Аликвоту гранулы массой 0,5 г смешивали с 40 мл 2 M NaOH и гидролизовали в потоке азота при постоянном перемешивании в течение 1 часа. Затем образец центрифугировали в течение 20 мин при 1400×g; супернатант подкисляли до pH 2 c 12 M HCl и затем подвергали процедуре экстракции, уже описанной для СВОБОДНОЙ фракции.
2.6.4. Анализ методом ВЭЖХ
По 20 мкл каждого экстракта вводили в систему ВЭЖХ (Jasco, Токио, Япония; насос PU-4180, MD-4015 PDA детектор, AS-4050 автосамплер). Стационарной фазой служила обращенно-фазовая колонка Agilent (Santa Clara, CA, USA) Zorbax Eclipse Plus C18 (внутренний диаметр 100 мм × 3 мм, 3.5 мкм).
Хроматографический метод анализа фенольных кислот был адаптирован Mattila и соавт. [30]. Градиентное элюирование проводили смесью кислого фосфатного буфера и ацетонитрила со скоростью 0,7 мл/мин. Для количественного определения анализируемого вещества использовали сигналы при 254, 280 и 329 нм. Значения извлечения фенольных кислот в образцах с добавками варьировались от 78,8 до 92,2% (RSD< 9,8%, n= 6).
2.7. Определение содержания каротиноидов методом ВЭЖХ
Образцы хлеба подвергали сублимационной сушке и тонкому измельчению в ножевой мельнице в течение 4 периодов по 30 секунд. Кроме того, дигесту in vitro перед экстракцией подвергали сублимационной сушке. Каротиноиды извлекали из образцов в соответствии со следующим протоколом: 5 г муки или сублимированного хлеба смешивали в течение 30 секунд в мельнице с 20 мл смеси этанол/вода (80/20, v/v), оставляли на 20 минут, затем снова перемешивали в течение 30 секунд и центрифугировали в течение 10 мин при 4000×g. Высушенные вымораживанием обработанные образцы (0,5 г) экстрагировали 2 мл смеси этанол/вода. Супернатант отфильтровывали и 20 мкл этого раствора вводили в систему ВЭЖХ (см. раздел 2.6). Метод анализа каротиноидов был адаптирован по Hidalgo и соавт. [31]. Для определения состава и градиента скорости потока использовали смесь воды и метанола/тетрагидрофурана (50/50, v/v). Сигнал при длине волны 450 нм использовался для количественных целей. Значения извлечения каротиноидов в пробах с добавками варьировались от 80,1 до 90,3% (RSD < 11,1%, n = 6).
2.8. Анализ поглощающей способности кислородных радикалов (ORAC)
Анализ проводили на экстрактах образцов хлеба с использованием мультиабразивного считывателя Victor X3 фирмы Perkin Elmer (Турку, Финляндия), по существу, как описано Moore и соавт. [29], с существенными изменениями. Для построения калибровочных кривых в каждую лунку вносили по 210 мкл 10 нМ флуоресцеина и 35 мкл троллокса в различных концентрациях (10-500 мкм) или 35 мкл 10 мм фосфатного буфера (чистый раствор). Для анализа пробы раствор Троллокса был заменен на 35 мкл соответственно разведенной пробы. Это инкубировали при 37°C в течение 5 мин, затем добавили 35 мкл 240 mM 2,2′- Азобис(2-метилпропионамидин) дигидрохлорида (AAPH). Интенсивность флуоресцентного излучения каждой лунки контролировали в течение 150 мин при 550 нм, возбуждая при 495 нм, при термостатировании при 28 °C. Эквиваленты троллокса (TE) были рассчитаны по относительной площади под кривой зависимости интенсивности излучения от времени.
2.9. Клеточная культура Caco-2 и пищевые добавки
Клетки Caco-2 хранили при температуре 37 ◦C, 5% CO2 в EBSS с добавлением 20% FBS, 100 ЕД/мл пенициллина, 100 мг/мл стрептомицина, 2 мм L-глютамина, 1% NEAA, 1 мм пирувата натрия и 0,4% фунгизона. Среду меняли каждые два дня. Примерно через 10 дней, при 80%-ном слиянии, клетки разделяли (1:2-1:4) в новой колбе (75 см2). Для проведения различных анализов клетки высевали в 24-луночные планшеты по 80 000 клеток на лунку в DMEM без фенолового красного, 10% FBS, 100 ЕД/мл пенициллина, 100 мг/мл стрептомицина, 2 мм L-глютамина, 1% NEAA. Через 7 дней в предварительных экспериментах некоторым клеткам добавляли продукт холостого расщепления (1:25 по объему) в течение 24 часов (B), в то время как другие клетки не получали никаких добавок (контрольные-C). В дальнейших экспериментах, после 6 ч добавления переваренных образцов (1:25 по объему), продукта холостого переваривания (1:25 по объему) или без добавления, воспаление индуцировали воздействием IL-1β (10 нг/мл) в течение 18 ч.
2.10. Количественное определение цитокинов в клетках Caco-2
Уровень провоспалительных IL-6 и IL-8 определяли в клеточных средах с помощью наборов AlphaLISA (Исследовательские наборы для иммуноанализа IL-6 и IL-8; Perkin Elmer Inc., Уолтем, Массачусетс, США) в соответствии с инструкциями производителя. Было использовано 96 микроячейковых пластин (96 пластин размером 1/2 дюйма от Perkin Elmer), которые считывались с помощью устройства для чтения пластин EnSpire™ от Perkin Elmer. Данные были интерполированы в стандартную кривую IL-6 и IL-8, и результаты были выражены в процентах от значения, полученного в контрольных, не стимулированных клетках (присвоено значение 100%).
Секрецию IL-6 клетками Caco-2 также косвенно оценивали с помощью клеток HEK-Blue™ IL-6 (Invivogen, Сан-Диего, Калифорния, США). Эти клетки были получены путем стабильной трансфекции клеток HEK293 человеческим геном IL-6R и STAT3-индуцируемым геном-репортером SEAP. При стимуляции IL-6 клетки HEK-Blue™ IL-6 запускают активацию STAT3 и последующую секрецию SEAP, которую можно контролировать с помощью колориметрического реактивного прибора QUANTI-Blue™ (Invivogen, Сан-Диего, Калифорния, США). Вкратце, клетки HEK выращивали в среде DMEM, 10% (v/v) FBS, 50 ед/мл пенициллина, 500 мкг/мл стрептомицина, 100 мкг/мл нормоцина, 2 мм L-глютамина и высевали в 96 многолуночных планшетах по 50 000 клеток на лунку. Клетки инкубировали при температуре 37°C и 5% CO2 течение 24 ч с непосредственным воздействием среды Caco-2 на клетки, выращенные в различных экспериментальных условиях. После инкубации активацию JAK-STAT-пути оценивали с помощью QUANTI-Blue™ в соответствии с инструкциями производителя.
2.11. Статистический анализ
Статистический анализ проводился с использованием однофакторного дисперсионного анализа (ANOVA) с использованием множественных сравнительных тестов Тьюки. Двухфакторный дисперсионный анализ (ANOVA) использовался для оценки влияния двух факторов (тип муки, ферментация) на уровни функциональных соединений. Содержание феруловой кислоты и активность ORACбыли сопоставлены с помощью корреляционного теста Пирсона.
3. Результаты и обсуждение
3.1. Содержание каротиноидов и фенольных кислот в однозернянке и пшеничной муке
Лютеин и зеаксантин были единственными каротиноидами, обнаруженными во всех образцах муки, причем первых было гораздо больше, чем вторых. Уровни каротиноидов в муке AF были в пределах ожидаемых значений [32] и в 7 раз выше, чем в муке SF, как сообщалось ранее [33]. В муке AF белого сорта содержание лютеина и зеаксантина было немного, но незначительно, ниже, чем в цельной. Это подтверждает высокий вклад эндосперма в содержание каротиноидов в семенах [34]. Существенных различий в содержании каротиноидов между испанской и польской пшеничной мукой обнаружено не было (рис. 1).
Что касается фенольных кислот, то наиболее распространенной была феруловая кислота, за которой следовала p-Кумаровая кислота (таблица 2). Как уже сообщалось, другие фенольные кислоты присутствовали в меньших количествах [6,35]. Как и ожидалось, феруловая кислота, которая связана с компонентами клеточной стенки, присутствовала в основном в форме IB. В AFсодержание феруловой кислоты в связанном виде было примерно в 4 раза выше, чем в белой муке SF; в отличие от AF, в целом виде мука имела самое высокое содержание феруловой кислоты. Разница между цельной мукой и белой мукой AF оказалась не такой очевидной, как ожидалось. Действительно, в последнем содержалось на 22% меньше феруловой кислоты, чем в первом. Это можно объяснить тем фактом, что белая мука AF – мука каменного помола, что позволило сохранить довольно высокое содержание клетчатки.
Что касается белой SF, то в испанских и польских образцах содержание феруловой кислоты было одинаковым. Уровни феруловой кислоты, определенные в нашей белой SF, соответствовали данным, опубликованным Mattila и соавт. [30], хотя другие авторы обнаружили более низкие уровни [11]. Эти расхождения, вероятно, связаны с различными условиями выращивания зерновых и различными методологическими процедурами анализа фенольных кислот. Содержание p-Кумаровой кислоты было значительно ниже, чем феруловой кислоты, и она была более равномерно распределена между тремя фракциями. Белая SF показала содержание свободной p-Кумаровой кислоты в 5 раз выше, чем в AF (Таблица 2).
3.2. Содержание каротиноидов и фенольных кислот в хлебе AF и SF, приготовленном различными способами ферментации
На основании результатов анализа на содержание каротиноидов и фенольных кислот в каждом сорте (AF и SF) были выбраны две разновидности муки с наибольшим содержанием каротиноидов и феруловой кислоты (цельная AF и SF 650 P) для приготовления хлеба обычным способом или на закваске.
Содержание каротиноидов в различных видах хлеба показано на рисунке 2. Основными обнаруженными каротиноидами были лютеин и зеаксантин, причем уровень зеаксантина был значительно ниже, чем уровень лютеина. Деградация каротиноидов в процессе выпечки хлеба в AF-хлебе происходила иначе, чем в SF-хлебе. В первых содержание лютеина и зеаксантина составляло от 58% до 76% от содержания в муке, в то время как во вторых сохранялся меньший процент каротиноидов (16-20%).
Leenhardt и соавт. [36] исследовали изменения содержания каротиноидов в процессе приготовления цельнозернового хлебы и хлеба из однозернянки и продемонстрировали, что стадия замешивания, помимо самого процесса выпечки, приводит к значительным потерям каротиноидов. Это связано с заметным поступлением кислорода в тесто на этой стадии, что способствует окислению полиненасыщенных жирных кислот, катализируемому липоксигеназой (LOX) с одновременным окислением каротиноидов.
Как сообщалось ранее [36], ферментация, по-видимому, очень слабо влияет на содержание каротиноидов, возможно, из-за потребления кислорода пекарскими дрожжами, которые предотвращают разрушение каротиноидов, опосредованное LOX. Зерна однозернянки обычно обладают гораздо более низкой активностью LOX, чем зерна пшеницы [37], и это может объяснить меньшую деградацию, наблюдаемую в хлебе AF.
Ферментация закваски существенно не повлияла на содержание каротиноидов по сравнению с обычной процедурой (рисунок 2). Таким образом, двусторонний дисперсионный анализ показывает, что изменения в уровнях каротиноидов в образцах хлеба были обусловлены только типом муки (р < 0,001), и ни процедура ферментации, ни взаимодействие между мукой и ферментацией не представляли собой существенных источников различий (данные не приведены).
В хлебобулочных изделиях, подвергнутых лабораторной ферментации, происходят некоторые изменения в функциональных соединениях и питательных веществах, но о каротиноидах имеется мало информации. Учитывая гораздо большую продолжительность брожения закваски, что приводит к более активному усвоению кислорода, в хлебобулочных изделиях, подвергнутых лабораторной ферментации, можно ожидать более выраженного разложения каротиноидов. В этом исследовании лаборатория, используемая для ферментации экспериментального хлеба, была выбрана исходя из их пригодности для повышения качества хлебобулочных изделий [38]. Сообщалось, что Lactobacillus plantarum 98a увеличивает поступление антиоксидантных соединений [39]. Возможно, что кислая среда и/или микробный метаболизм каким-то образом препятствовали активности ферментов, разрушающих каротиноиды. Кроме того, производство новых биологически активных соединений в продуктах на основе закваски может быть обусловлено специфическими бактериальными синтетическими путями. Например, было показано, что штаммы Lactobacillus plantarum обладают темно-желтой пигментацией при культивировании в виде изолированных колоний [40], и этот вид может синтезировать желтый каротиноид С30 4,4- диапонейроспорен [41].
Во всех видах хлеба феруловая кислота была наиболее распространенной фенольной кислотой, и основной вклад в ее содержание вновь внесла фракция IB (>90% от общего количества) (рисунок 3). Эффект ферментации закваски был различным в зависимости от типа используемой муки: в хлебобулочных изделиях AF лабораторная ферментация в значительной степени поддерживала исходный уровень феруловой кислоты (11 и 17 мкг/г в хлебобулочных изделиях против 18 мкг/г в муке), в то же время приводя к значительному снижению связанной фракции. И наоборот, в хлебе SF не было обнаружено существенных изменений ни в SC, ни в IB форме феруловой кислоты, в то время как в хлебе, приготовленном путем брожения на закваске с использованием AF в качестве муки для закваски, наблюдалось значительное увеличение содержания свободной феруловой кислоты.
Таким образом, двусторонний аналитический анализ показал, что оба фактора, тип муки и ферментация, а также их взаимодействие, были значимыми при определении различий в содержании феруловой кислоты в различных видах хлеба (р < 0,05 для СВОБОДНОЙ фракции и р < 0,01 для других фракций). Ферментация внесла основной вклад в изменение общего содержания растворимой феруловой кислоты (свободная + SC, р < 0,001). Общее содержание феруловой кислоты в муке и соответствующих хлебобулочных изделиях было одинаковым, что позволяет предположить, что в процессе приготовления хлеба фенольные кислоты не изменялись в значительной степени. Сообщалось, что количество свободных фенольных соединений и феруловой кислоты увеличивается при ферментации ржаных отрубей с использованием пекарских дрожжей [42] и при ферментации в лабораторных условиях ржаной, овсяной и ячменной муки грубого помола [43,44]. Это было связано с использованием заквасочных культур, обладающих ферулоилэстеразной активностью. Однако сообщалось, что влияние культур на содержание фенольной кислоты или ее отсутствие в высокой степени зависит от штамма по разным причинам [44,45]. Внутренняя активность ферулоилэстеразы в злаках также способствует метаболизму фенолов [19], и сообщалось о различном эффекте в ячневой и овсяной крупяной муке, ферментированной одним и тем же штаммом [44]. Таким образом, наблюдаемое снижение содержания феруловой кислоты IB в хлебе AF, сопровождающееся значительным увеличением доли SC, может быть объяснено связыванием свободной феруловой кислоты, образующейся в результате гидролиза связанной формы, с молекулами низкой молекулярной массы (такими как небольшие пептиды и аминокислоты), образующимися в процессе ферментации [46]. Различный характер превращения фенольных кислот в хлебе AF и SF может быть обусловлен различной активностью эндогенной ферулоилэстеразы зерновых культур, которая зависит от компонента отрубей AF, активируемого во время ферментации.
Анализы ORAC (измеряющие антиоксидантную способность различных веществ) показывают, что фракция IB обладала наибольшей активностью, в то время как СВОБОДНАЯ фракция и SC обладали сходной и более низкой активностью (рис. 4). Тем не менее, различия между IB, SC и СВОБОДНЫМИ фракциями в активности ORAC были значительно ниже, чем различия в содержании феруловой кислоты, которая присутствовала во фракции IB на уровнях, в 200 раз превышающих уровни SC (рисунки 3 и 4). Корреляцию между содержанием ORAC и феруловой кислоты во фракциях SC и IB оценивали с помощью корреляционного теста Пирсона, и была обнаружена лишь умеренная корреляция (значения r Пирсона = 0,71 и 0,60 соответственно). Это неудивительно и может быть объяснено тем фактом, что экстракты из этих фракций, скорее всего, представляют собой сложные смеси, содержащие различные классы молекул, взаимодействующих друг с другом посредством синергизма и/или антагонизма, что может привести к непредсказуемой антиоксидантной активности [47]. Более того, различные классы соединений в экстрактах могут иметь механизм реакции, отличный от переноса атомов водорода, на котором основан анализ ORAC. Как тип муки, так и процедура ферментации представляли собой значительный источник различий в активности по удалению радикалов во фракциях SC и IB, в то время как взаимодействие между этими двумя факторами было незначительным (данные не приведены).
3.3. Каротиноиды и фенольные кислоты в образцах, обработанных In Vitro
Каротиноиды и фенольные кислоты в растворимой фракции, полученной после переваривания образцов хлеба in vitro, представляют собой биодоступные соединения, т.е. количество соединений, высвобождаемых в процессе переваривания и доступных для усвоения. Как видно из таблицы 3, процентное содержание биодоступных лютеина и зеаксантина было довольно высоким, достигая в некоторых образцах более 60%. Это согласуется с предыдущими исследованиями, показавшими более чем 65%-ную биодоступность каротиноидов [48,49].
Хлеб, приготовленный путем ферментации на закваске, показал более низкую биодоступность лютеина и зеаксантина по сравнению с хлебом, сброженным обычными дрожжами, что позволяет предположить, что извлечение лютеина и зеаксантина из хлебной массы происходит в разной степени в зависимости от процедуры ферментации, или что при переходе желудочно-кишечного тракта может происходить более интенсивное разложение каротиноидов. Таким образом, возможно, что ферментация закваски за счет увеличения количества пищевых волокон за счет большей растворимости арабиноксиланов [50] повлияла на удаление каротиноидов из хлебной массы, о чем уже сообщали другие авторы [51].
Биодоступность феруловой кислоты была очень низкой (менее 1% в хлебе AF; таблица 3), что согласуется с предыдущими отчетами [24] и с тем фактом, что большая ее часть была связана с арабиноксиланами и другими полисахаридами клеточной стенки, которые могут препятствовать перевариванию в верхних отделах желудочно-кишечного тракта [52]. Феруловая кислота была обнаружена только в хлебе AF, в то время как ее содержание в хлебе SF было ниже предела обнаружения, без существенных различий между обычным хлебом и хлебом на закваске (таблица 3). Эти результаты подтверждают гипотезу о том, что расщепление сложноэфирной связи в гидроксициннаматах происходит в основном в толстой кишке и опосредуется бактериальными ферментами [52,53].
3.4. Противовоспалительный эффект хлеба, переваренного In Vitro
Предполагаемый противовоспалительный эффект однозернянки оценивали, сравнивая растворимую фракцию четырех видов ферментированного хлеба, т.е. хлеба из однозернянки, приготовленного с использованием как обычной ферментации, так и ферментации на закваске (AFCF и AFSAF), и пшеничного хлеба, приготовленного с использованием тех же видов ферментации (SFCF и SFSAF). К клеткам Caco-2 добавляли растворимую фракцию переваренного хлеба, а противовоспалительный эффект оценивали путем измерения секреции IL-6 и IL-8, которые, как сообщается, являются основными факторами воспаления [54]. Поскольку было показано, что желчные кислоты запускают окисление [55] и играют определенную роль в патогенезе воспаления кишечника [56], в предварительных экспериментах к клеткам добавляли продукт холостого переваривания, то есть переваривания in vitro без пищи (В). В дальнейших экспериментах клетки Caco-2 подвергали воздействию воспалительного стимула (обработка IL-1β), а затем добавляли к ним различные переваренные хлебцы или пустую дижесту. Во всех экспериментах результаты, полученные в клетках с добавками, сравнивались с парно подобранными клетками без добавок (US).
В исходном состоянии добавление продукта холостого переваривания не изменяло секрецию IL-6 и IL-8, которая была значительно повышена во всех группах клеток при обработке IL-1β (рис. 5).
Уровень IL-6 повышался в клетках US в большей степени, чем в клетках с добавкой, за исключением SFSAF, в то время как увеличение секреции IL-8 было одинаковым во всех клетках.
В отличие от других исследований, в которых сообщалось о снижении концентрации IL-8 при обработке непереваренной или переваренной черной морковью, кожурой и выжимками, богатыми полифенолами и каротиноидами, в клетках Caco-2 [57,58], мы не наблюдали каких-либо изменений в секреции IL-8 в ответ на воспалительный стимул. Это можно объяснить различной концентрацией добавляемых биоактивных веществ. Фактически, количество фенольных кислот, добавляемых в клетки в нашем исследовании (не более 30 нМ в эквиваленте феруловой кислоты), было по меньшей мере в 100 раз меньше, чем в [58]. Высокая концентрация фенольной кислоты не похожа на физиологическую, базальную ситуацию. Хотя они были обнаружены в плазме крови человека после острого или хронического диетического вмешательства [59,60] и возможно, что они могут оказывать противовоспалительное действие, оценка их эффективности выходила за рамки настоящего исследования.
Секреция IL-6 в ответ на воспалительные процессы была снижена при добавлении как в готовую пищу, так и в переваренный хлеб. Таким образом, было трудно провести различие между противовоспалительным действием желчных кислот [61] и вкладом биологически активных веществ в хлеб. Поскольку IL-6 играет ключевую роль в индукции и поддержании воспаления в кишечнике посредством активации пути JAK-STAT [62], для дальнейшего определения эффекта различных сортов хлеба активация пути JAK-STAT под действием IL-6, продуцируемого клетками Caco-2, была проанализирована на HEK-Blue™ IL-6 клетках, которые подвергали воздействию контрольной среды и дополняли клетками Caco-2 (рис. 6).
Активация JAK-STAT не была обнаружена в исходном состоянии, в то время как в клетках Caco-2, обработанных IL-1β, она в значительной степени индуцировалась всеми средами. Результаты на клетках HEK-Blue™ подтвердили более низкую выработку IL-6 в дополненных клетках, чем в клетках, обработанных US, при воспалительном воздействии. При сравнении воспаленных клеток с дополненными клетками, активация JAK-STAT была значительно ниже в AFCF, чем в клетках группы В. Таким образом, хотя противовоспалительный эффект, по-видимому, был обусловлен в основном пищеварительными жидкостями, биологически активные вещества в AFCF внесли свой вклад в конечный эффект.
4. Выводы
Инновационная разработка функциональных хлебобулочных изделий требует знаний о влиянии рецептур и технологических процессов на содержание полезных для здоровья компонентов в готовых изделиях. Хотя недавние исследования показывают, что древние злаки могут оказывать положительное влияние на сердечно-сосудистую систему [63] и уменьшать воспаление [64] и, следовательно, риск заболеваний, связанных с воспалением, таких как болезнь раздраженного кишечника [65], исследования однозернянки все еще относительно недостаточно развиты.
Насколько известно авторам, это исследование представляет собой первую комплексную оценку потенциальной пользы для здоровья хлеба на основе однозернянки по сравнению с хлебом на основе пшеницы, учитывая химические характеристики муки, влияние различных процессов ферментации, влияние переваривания на биодоступность биологически активных соединений и биологический эффект переваренного хлеба на культивируемые клетки кишечника.
Наши результаты подтверждают более высокий уровень каротиноидов в однозернянке, чем в современной пшенице, что, по крайней мере, частично объясняет полезные свойства однозернянки для здоровья. Кроме того, использование ферментации на закваске, по-видимому, позволяет сохранить каротиноиды в конечном продукте, несмотря на более длительное время, необходимое для обработки. Также произошли значительные изменения в составе фенольных кислот хлеба, в частности, изменилось их распределение, что явно указывает на специфику для каждого вида злака. Более того, результаты экспериментов по ферментации in vitro показали, что ферментация на закваске влияет на биодоступность каротиноидов, при этом они лучше сохраняются в хлебе микробиологической закваски по сравнению с хлебом, заквашенным на дрожжах, что, возможно, связано с более высокой растворимостью волокон, опосредованной молочнокислыми бактериями. Хотя предполагаемый противовоспалительный эффект хлеба в клетках Caco-2 был замаскирован воздействием пищеварительной жидкости, эксперименты с использованием клеток HEK-Blue™ IL-6 показали защитный эффект хлеба из однозернянки, приготовленного обычным способом ферментации. Примечательно, что в этой модельной системе не учитывался вклад кишечной микробиоты в трансформацию полифенолов, и это могло бы объяснить более низкую эффективность однозернянки в борьбе с воспалением, о которой уже сообщалось для других древних злаков в исследованиях in vivo [64,65].
Несмотря на это, представленные здесь результаты подтверждают, что однозернянка является хорошим кандидатом для производства хлебобулочных изделий с улучшенными питательными свойствами.
Благодарности: Эта работа была профинансирована Седьмой рамочной программой Европейского союза (European Union’s Seventh Framework Program), управляемой REA-Research Executive Agency http://ec.europa.eu/research/rea (FP7/2007-2013) в рамках грантового соглашения n. 606476. Авторы выражают благодарность Massimo Fiorani (Prometeo s.r.l, Urbino, Italy за поставку муки из однозернянки, Maria Luisa Llin Albinana (INDESPAN—Investigacion y Desarrollo Panadero, S.L, Valencia, Spain) и Liliana Rehrer Rychel and Radek Rychel (VINI, Rogoznik, Poland) за испанскую и польскую пшеничную муку, соответственно.
Вклад авторов: F.A. и R.M. провели анализ каротиноидов и фенольных кислот; M.D.N. провели анализ клеток Caco-2; A.G., A.B., F.A. и R.M. разработали исследование и написали статью. D.L.T.S. и A.G. предоставили штаммы L.A.B. и приготовили экспериментальный хлеб. B.V., E.G., M.P.V., L.T.-C. провели обработку in vitro и эксперименты с клетками HEK. Все авторы внесли значительный вклад в написание рукописи.
Конфликт интересов: Авторы заявляют об отсутствии конфликта интересов. Спонсоры не принимали участия в разработке проекта исследования, сборе, анализе или интерпретации данных, написании рукописи и принятии решения о публикации результатов.
Источники
1. Zong, G.; Gao, A.; Hu, F.B.; Sun, Q. Whole grain intake and mortality from all causes, cardiovascular disease, and cancer: A meta-analysis of prospective cohort studies. Circulation 2016, 133, 2370–2380. [CrossRef] [PubMed]
2. Aune, D.; Keum, N.; Giovannucci, E.; Fadnes, L.T.; Boffetta, P.; Greenwood, D.C.; Tonstad, S.; Vatten, L.J.; Riboli, E.; Norat, T. Whole grain consumption and risk of cardiovascular disease, cancer, and all cause and cause specific mortality: Systematic review and dose-response meta-analysis of prospective studies. BMJ 2016, 353, i2716. [CrossRef] [PubMed]
3. Ye, E.Q.; Chacko, S.A.; Chou, E.L.; Kugizaki, M.; Liu, S. Greater whole-grain intake is associated with lower risk of type 2 diabetes, cardiovascular disease, and weight gain. J. Nutr. 2012, 142, 1304–1313. [CrossRef] [PubMed]
4. Kyrø, C.; Skeie, G.; Loft, S.; Landberg, R.; Christensen, J.; Lund, E.; Nilsson, L.M.; Palmqvist, R.; Tjønneland, A.; Olsen, A. Intake of whole grains from different cereal and food sources and incidence of colorectal cancer in the Scandinavian HELGA cohort. Cancer Causes Control 2013, 24, 1363–1374. [CrossRef] [PubMed]
5. Adom, K.K.; Liu, R.H. Antioxidant activity of grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [CrossRef]
[PubMed]
6. Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemical profiles and antioxidant activity of wheat varieties.
J. Agric. Food Chem. 2003, 51, 7825–7834. [CrossRef] [PubMed]
7. Abdel-Aal, E.S.M.; Young, J.C.; Rabalski, I.; Hucl, P.; Frégeau-Reid, J. Identification and quantification of seed carotenoids in selected wheat species. J. Agric. Food Chem. 2007, 55, 787–794. [CrossRef] [PubMed]
8. Perez-Jimenez, J.; Neveu, V.; Vos, F.; Scalbert, A. Systematic analysis of the content of 502 polyphenols in 452 foods and beverages: An application of the Phenol−Explorer database. J. Agric. Food Chem. 2010, 58, 4959–4969. [CrossRef] [PubMed]
9. Abdel-Aal, E.S.M.; Young, J.C.; Wood, P.J.; Rabalski, I.; Hucl, P.; Fregeau-Reid, J. Einkorn: A potential candidate for developing high lutein wheat. Cereal Chem. 2002, 79, 455–457. [CrossRef]
10. Humphries, J.M.; Khachik, F. Distribution of lutein, zeaxanthin and related geometrical isomers in fruit, vegetables, wheat and pasta products. J. Agric. Food Chem. 2003, 51, 1322–1327. [CrossRef] [PubMed]
11. Sosulski, F.; Krygier, K.; Hogge, L. Free, esterified, and insoluble-bound phenolic acids. 3. Composition of phenolic acids in cereal and potato flour. J. Agric. Food Chem. 1982, 30, 337–340. [CrossRef]
12. Poutanen, K.; Shepherd, R.; Shewry, P.R.; Delcour, J.A.; Bjorck, I.; Van Der Kamp, J.W. Beyond whole grain: The European HEALTH GRAIN project aims at healthier cereal foods. Cereal Foods World 2008, 53, 32–35. [CrossRef]
13. Slavin, J. Why whole grains are protective: Biological mechanisms. Proc. Nutr. Soc. 2003, 62, 129–134. [CrossRef] [PubMed]
14. Bordoni, A.; Danesi, F.; Di Nunzio, M.; Taccari, A.; Valli, V. Ancient wheat and health: A legend or the reality? A review on KAMUT khorasan wheat. Int. J. Food Sci. Nutr. 2016, 28, 1–9. [CrossRef] [PubMed]
15. Hidalgo, A.; Brandolini, A. Nutritional properties of einkorn wheat (Triticum monococcum L.). J. Sci. Food Agric.
2014, 94, 601–612. [CrossRef] [PubMed]
16. Molberg, Ø.; Uhlen, A.K.; Jensen, T.; Flæte, N.S.; Fleckenstein, B.; Arentz-Hansen, H.; Raki, M.; Lundin, K.E.A.; Sollid, L.M. Mapping of gluten T-cell epitopes in the bread wheat ancestors: Implication for celiac disease. Gastroenterology 2005, 128, 393–401. [CrossRef] [PubMed]
17. Sánchez-Pardo, M.E.; Blancas-Nápoles, J.A.; Vázquez-Landaverde, P.A.; Nari, A.; Taglieri, I.; Ortiz-Moreno, A.;
Mayorga-Reyes, L.; Sanmartin, C.; Bermúdez-Humarán, L.G.; Torres-Maravilla, E. The use of Mexican xaxtle as leavening agent in Italian straight dough bread making to produce pulque bread. Agrochimica 2016, 60, 329–342.
18. Venturi, F.; Sanmartin, C.; Taglieri, I.; Nari, A.; Andrich, G.; Zinnai, A. Effect of the baking process on artisanal sourdough bread-making: A technological and sensory evaluation. Agrochimica 2016, 60, 222–234.
19. Ganzle, M.G. Enzymatic and bacterial conversions during sourdough fermentation. Food Microbiol. 2014, 37,
20. Gobbetti, M.; Rizzello, G.C.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [CrossRef] [PubMed]
21. Venturi, F.; Sanmartin, C.; Taglieri, I.; Nari, A.; Andrich, G.; Terzuoli, E.; Donnini, S.; Nicolella, C.; Zinnai, A. Development of phenol-enriched olive oil with phenolic compounds extracted from wastewater produced by physical refining. Nutrients 2017, 9, 916. [CrossRef] [PubMed]
22. Turpin, W.; Renaud, C.; Avallone, S.; Hammoumi, A.; Guyot, J.-P.; Humblot, C. PCR of crtNM combined with analytical biochemistry: An efficient way to identify carotenoid producing lactic acid bacteria. Syst. Appl. Microbiol. 2016, 39, 115–121. [CrossRef] [PubMed]
23. Reboul, E.; Richelle, M.; Perrot, E.; Smoulins-Malezet, C.; Pirisi, V.; Borel, P. Bioaccessibility of carotenoids and vitamin E from their main dietary sources. J. Agric. Food Chem. 2006, 54, 8749–8755. [CrossRef] [PubMed]
24. Anson, N.M.; van den Berg, R.; Havenaar, R.; Bast, A.; Haenen, G.R.M.M. Bioavailability of ferulic acid is determined by its bioaccessibility. J. Cereal Sci. 2009, 49, 296–300. [CrossRef]
25. Viadel Crespo, B.; Rivera Patiño, J.D.; Navarro Fayos, M.T.; Tenllado Llavador, I.; Carreres Malonda, J.E.; García Reverter, J.; Blasco Piquer, M.; Subirats Huerta, S. Equipo modular de digestión in vitro. Patent ES2361983B1, 19 Apirl 2012.
26. Marteau, P.; Flourié, B.; Pochart, P.; Chastang, C.; Desjeux, J.F.; Rambaud, J.C. Role of the microbial lactose EC 3.2.123, activity from yoghurt on the intestinal absorption of lactose: An in vivo study in lactose-deficient humans. Br. J. Nutr. 1990, 64, 71–79. [CrossRef] [PubMed]
27. Minekus, M.; Marteau, P.; Havenaar, R.; Huis Veld, J.H.J. A multicompartmental dynamic computer-controlled model simulating the stomach and the small intestine. ATLA 1995, 23, 197–209.
28. Bengtsson, A.; Larsson, M.; Svanberg, U. In vitro bioaccessibility of β-carotene from heat-processed
orange-fleshed sweet potato. J. Agric. Food Chem. 2009, 57, 9693–9698. [CrossRef] [PubMed]
29. Moore, J.; Hao, Z.; Zhou, K.; Luther, M.; Costa, J.; Yu, L. Carotenoid, tocopherol, phenolic acid, and antioxidant properties of Maryland-grown soft wheat. J. Agric. Food Chem. 2005, 53, 6649–6657. [CrossRef] [PubMed]
30. Mattila, P.; Pihlava, J.M.; Hellström, J. Contents of phenolic acids, alkyl- and alkenylresorcinols, and avenanthramides in commercial grain products. J. Agric. Food Sci. 2005, 53, 8290–8295. [CrossRef] [PubMed]
31. Hidalgo, A.; Brandolini, A.; Pompei, C.; Piscozzi, R. Carotenoids and tocols of einkorn wheat (Triticum monococcum ssp. monococcum L.). J. Cereal Sci. 2006, 44, 182–193. [CrossRef]
32. Brandolini, A.; Castoldi, P.; Plizzari, L.; Hidalgo, A. Phenolic acid composition, total polyphenols content and antioxidant activity of Triticum monococcum, Triticum turgidum and Triticum aestivum: A two-years evaluation.
J. Cereal Sci. 2013, 58, 123–131. [CrossRef]
33. Abdel-Aal, E.S.M.; Rabalski, I. Bioactive compounds and their antioxidant capacity in selected primitive and modern wheat species. Open Agric. J. 2008, 2, 7–14. [CrossRef]
34. Hidalgo, A.; Brandolini, A. Protein, ash, lutein and tocols distribution in einkorn (Triticum monococcum L.
subsp. Monococcum) seed fractions. Food Chem. 2008, 107, 444–448. [CrossRef]
35. Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemicals and antioxidant activity of milled fractions of different wheat varieties. J. Agric. Food Chem. 2005, 53, 2297–2306. [CrossRef] [PubMed]
36. Leenhardt, F.; Lyan, B.; Rock, E.; Boussard, A.; Potus, J.; Chanliaud, E.; Remesy, C. Wheat lipoxygenase activity induces greater loss of carotenoids than Vitamin E during breadmaking. J. Agric. Food Chem. 2006, 54, 1710–1715. [CrossRef] [PubMed]
37. Leenhardt, F.; Lyan, B.; Rock, E.; Boussard, A.; Potus, J.; Chanliaud, E.; Remesy, C. Genetic variability of carotenoid concentration, and lipoxygenase and peroxidase activities among cultivated wheat species and bread wheat varieties. Eur. J. Agron. 2006, 25, 170–176. [CrossRef]
38. Cevoli, C.; Gianotti, A.; Troncoso, R.; Fabbri, A. Quality evaluation by physical tests of a traditional Italian flat bread Piadina during storage and shelf-life improvement with sourdough and enzymes. Eur. Food Res. Technol. 2015, 240, 1081–1089. [CrossRef]
39. Ferri, M.; Serrazanetti, D.I.; Tassoni, A.; Baldissarri, M.; Gianotti, A. Improving functional and technological profile of cereal fermented foods by Lactobacillus plantarum strains selected via a metabolomics approach. Food Res. Int. 2016, 89, 1095–1105. [CrossRef]
40. Garrido-Fernandez, J.; Maldonado-Barragan, A.; Caballero-Guerrero, B.; Hornero-Mendez, D.; Ruiz-Barba, J.L. Carotenoid production in Lactobacillus plantarum. Int. J. Food Microbiol. 2010, 140, 34–39. [CrossRef] [PubMed]
41. Breithaupt, D.E.; Schwack, W.; Wolf, G.; Hammes, W.P. Characterization of the triterpenoid diaponeurosporene and its isomers in food-associated bacteria. Eur. Food Res. Technol. 2001, 213, 231–233. [CrossRef]
42. Katina, K.; Laitila, A.; Jovonen, R.; Liukkonen, K.-H.; Kariluoto, S.; Piironen, V.; Landgerg, R.; Åman, P.; Poutanen, K. Bran fermentation as a means to enhance technological properties and bioactivity of rye. Food Microbiol. 2007, 24, 175–186. [CrossRef] [PubMed]
43. Boskov Hansen, H.; Andreasen, M.G.; Nielsen, M.M.; Larsen, L.M.; Bach Knudsen, K.E.; Meyer, A.S.; Christensen, L.P.; Hansen, Å. Changes in dietary fibre, phenolic acids and activity of endogenous enzymes during rye bread-making. Eur. Food Res. Technol. 2002, 214, 33–42. [CrossRef]
44. Hole, A.S.; Rud, I.; Grimmer, S.; Sigl, S.; Narvhus, J.; Sahlstrøm, S. Improved bioavailability of dietary phenolics in whole grain barley and oat groat following fermentation with probiotic Lactobacillus acidophilus, Lactobacillus johnsonii, and Lactobacillus reuteri. J. Agric. Food Chem. 2012, 60, 6369–6375. [CrossRef] [PubMed]
45. Svensson, L.; Sekwati-Monang, B.; Lutz, D.L.; Schieber, A.; Ganzle, M.G. Phenolic acids and flavonoids in nonfermented and fermented red sorghum Sorghum bicolor L., Moench. J. Agric. Food Chem. 2010, 58, 9214–9220. [CrossRef] [PubMed]
46. Gobbetti, M.; De Angelis, M.; Corsetti, A.; Di Cagno, R. Biochemistry and physiology of sourdough lactic acid bacteria. Trends Food Sci. Technol. 2005, 16, 57–69. [CrossRef]
47. Makris, D.P.; Boskou, G.; Andrikopoulos, N.K. Polyphenolic content and in vitro antioxidant characteristics of wine industry and other agri-food solid waste extracts. J. Food Compos. Anal. 2007, 20, 125–132. [CrossRef]
48. Read, A.; Wright, A.; Abdel-Aal, E.S.M. In vitro bioaccessibility and monolayer uptake of lutein from wholegrain baked foods. Food Chem. 2015, 174, 263–269. [CrossRef] [PubMed]
49. Courraud, J.; Berger, J.; Cristol, J.P.; Avallone, S. Stability and bioaccessibility of different forms of carotenoids and vitamin A during in vitro digestion. Food Chem. 2013, 136, 871–877. [CrossRef] [PubMed]
50. Katina, K.; Juvonen, R.; Laitila, A.; Flander, L.; Nordlund, E.; Kariluoto, S.; Piironen, V.; Poutanen, K. Fermented wheat bran as a functional ingredient in baking. Cereal Chem. 2012, 89, 126–134. [CrossRef]
51. O’Connell, O.; Ryan, L.; O’Sullivan, L.; Aherne-Bruce, S.A.; O’Brien, N.M. Carotenoid micellarization varies greatly between individual and mixed vegetables with or without the addition of fat or fiber. Int. J. Vitam. Nutr. Res. 2008, 78, 238–246. [CrossRef] [PubMed]
52. Zhao, Z.; Egashira, Y.; Sanada, H. Digestion and absorption of ferulic acid sugar esters in rat gastrointestinal tract. J. Agric. Food Chem. 2003, 51, 5034–5039. [CrossRef] [PubMed]
53. Kroon, P.A.; Faulds, C.B.; Ryden, P.; Robertson, J.A.; Williamson, G. Release of covalently bound ferulic acid from fiber in the human colon. J. Agric. Food Chem. 1997, 45, 661–667. [CrossRef]
54. Laveti, D.; Kumar, M.; Hemalatha, R.; Sistla, R.; Naidu, V.G.; Talla, V.; Verma, V.; Kaur, N.; Nagpal, R. Anti-inflammatory treatments for chronic diseases: A review. Inflamm. All. Drug Targets 2013, 12, 349–361. [CrossRef] [PubMed]
55. Araki, Y.; Katoh, T.; Ogawa, A.; Bamba, S.; Andoh, A.; Koyama, S.; Fujiyama, Y.; Bamba, T. Bile acid modulates transepithelial permeability via the generation of reactive oxygen species in the Caco-2 cell line. Free Radic. Biol. Med. 2005, 39, 769–780. [CrossRef] [PubMed]
56. Di Toro, R.; Campana, G.; Murari, G.; Spampinato, S. Effects of specific bile acids on c-fos messenger RNA levels in human colon carcinoma Caco-2 cells. Eur. J. Pharm. Sci. 2000, 11, 291–298. [CrossRef]
57. Kaulmann, A.; Bohn, T. Carotenoids, inflammation, and oxidative stress—Implications of cellular signaling pathways and relation to chronic disease prevention. Nutr. Res. 2014, 34, 907–929. [CrossRef] [PubMed]
58. Kamiloglu, S.; Grootaert, C.; Capanoglu, E.; Ozkan, C.; Smagghe, G.; Raes, K.; Van Camp, J. Anti-inflammatory potential of black carrot (Daucus carota L.) polyphenols in a co-culture model of intestinal Caco-2 and endothelial EA.hy926 cells. Mol. Nutr. Food Res. 2017, 61. [CrossRef] [PubMed]
59. Marmet, C.; Actis-Goretta, L.; Renouf, M.; Giuffrida, F. Quantification of phenolic acids and their methylates, glucuronides, sulfates and lactones metabolites in human plasma by LC-MS/MS after oral ingestion of soluble coffee. J. Pharm. Biomed. Anal. 2014, 88, 617–625. [CrossRef] [PubMed]
60. McKay, D.L.; Chen, C.Y.; Zampariello, C.A.; Blumberg, J.B. Flavonoids and phenolic acids from cranberry juice are bioavailable and bioactive in healthy older adults. Food Chem. 2015, 168, 233–240. [CrossRef] [PubMed]
61. Ward, J.B.J.; Lajczak, N.K.; Kelly, O.B.; O’Dwyer, A.M.; Giddam, A.K.; Ní Gabhann, J.; Franco, P.; Tambuwala, M.M.; Jefferies, C.A.; Keely, S.; et al. Ursodeoxycholic acid and lithocholic acid exert anti-inflammatory actions in the colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G550–G558. [CrossRef] [PubMed]
62. Fantini, M.C.; Pallone, F. Cytokines: From gut inflammation to colorectal cancer. Curr. Drug Targets. 2008, 9, 375–380. [CrossRef] [PubMed]
63. Sereni, A.; Cesari, F.; Gori, A.M.; Maggini, N.; Marcucci, R.; Casini, A.; Sofi, F. Cardiovascular benefits from ancient grain bread consumption: Findings from a double-blinded randomized crossover intervention trial. Int. J. Food Sci. Nutr. 2017, 68, 97–103. [CrossRef] [PubMed]
64. Benedetti, S.; Primiterra, M.; Tagliamonte, M.C.; Carnevali, A.; Gianotti, A.; Bordoni, A.; Canestrari, F. Counteraction of oxidative damage in the rat liver by an ancient grain (Kamut brand khorasan wheat). Nutrition 2012, 28, 436–441. [CrossRef] [PubMed]
65. Sofi, F.; Whittaker, A.; Gori, A.M.; Cesari, F.; Surrenti, E.; Abbate, R.; Gensini, G.F.; Benedettelli, S.; Casini, A. Effect of Triticum turgidum subsp. turanicum wheat on irritable bowel syndrome: A double-blinded randomised dietary intervention trial. Br. J. Nutr. 2014, 111, 1992–1999. [CrossRef] [PubMed]
ОБ АВТОРАХ:
Fabiana Antognoni 1, Roberto Mandrioli 1, Alessandra Bordoni 2, , Mattia Di Nunzio 2,*, Blanca Viadel 3, Elisa Gallego 3, María Paz Villalba 3, Lidia Tomás-Cobos 3, Danielle Laure Taneyo Saa 2 and Andrea Gianotti 2
- Department for Life Quality Studies, University of Bologna, Corso Augusto 237, 47921 Rimini, Italy; fabiana.antognoni@unibo.it (F.A.); roberto.mandrioli@unibo.it (R.M.)
- Department of Agri-Food Sciences and Technologies, University of Bologna, Piazza Goidanich 60, 47521 Cesena, Italy; alessandra.bordoni@unibo.it (A.B.); danielle.taneyosaa2@unibo.it (D.L.T.S.); andrea.gianotti@unibo.it (A.G.)
- Department of New Products and Department of Bioassays, AINIA Technological Centre,c/Benjamín Franklin 5-11, Paterna, 46980 Valencia, Spain; bviadel@ainia.es (B.V.); egallego@ainia.es (E.G.); mpvillalba@ainia.es (M.P.V.); ltomas@ainia.es (L.T.-C.)
* Correspondence: mattia.dinunzio@unibo.it; Tel.: +39-0547-338-957
Nutrients Получено: 11 октября 2017 г.; Принято к рассмотрению: 9 ноября 2017 г.; Опубликовано: 11 ноября 2017 г.