Доброго времени суток дорогие мои подписчики, гости и читатели канала. Благодарю за вашу активность в предыдущих статьях. Это очень стимулирует автора к дальнейшему ведению канала. Было задано не мало интересных вопросов. И в этой статье я постараюсь дать на них ответы, тем самым подведя итоги по проартикулятам.
Вступление.
Эдиакарский период (Венд) - одна и самых загадочных эпох на Земле. Удивительно то, что чем больше мы узнаем об этом участке времени, тем еще больше вопросов. Мы познакомились с огромным количеством самых разных представителей фауны. Еще больше предстоит познакомиться. Материала, добытого учеными из разных уголков Земли, хватит мне еще на десяток (а то и больше) статей.
Один из вопросов, от кого произошли сами проартикуляты и чьими предками они же являются? Или это абсолютно тупиковая ветвь эволюции, так и не давшее "потомства" на эволюционном древе? Очередной "Черновик Бога"?
Но от куда в последствии появятся стрекающие, кишечнополостные, членистоногие, кольчатые черви, моллюски? От кого? Все эти вопросы очень важны. Вендобионты могут раскрыть и эту тайну.
Океан в эпоху Эдиакария.
На дне океана систематически наростали бактериальные маты. Кислотность Мирового Океана уже была значительно ниже, чем, скажем, во время Статерия или Риасия.
Но океан был еще весьма солёным. Намного более соленым, чем современный океан. Его самой доминирующей формой жизни по прежнему оставались бактерии и одноклеточные организмы. Именно они строили те самые бактериальные маты. Слой за слоем. А в самой толще океана мириады фито- и зоопланктона. Вода в океанах была мутной, по цвету напоминала раствор воды с молоком. И конечно солнечные лучи плохо рассеивались в такой воде. Мест для фотосинтеза было крайне мало.
По мнению старшего научного сотрудника ГИН РАН Антона Владимировича Колесникова, эдиакарская биота населяла глубины в пределах фотической зоны (освещаемой Солнцем верхней толщи воды). В современных морях и океанах глубина фотической зоны достигает 200 м ― до такой глубины возможен фотосинтез. Конечно, еще раз уточню, что это в современном Океане. В эдиакарскую эпоху глубина фотической зоны была значительно иной. Вероятно представляя собой почти самую кромку мелководий. Ведь естественных фильтраторов 550-600 миллионов лет назад еще небыло.
Тут стоит вспомнить о несправедливо подзабытых мною многоклеточных животных типа петалонам.
Высказывалось предположение, что данный тип селился в более глубоководной зоне Океана, с некоторым подводным течением, что способствовало размножению этих организмов. Напомню, что проследив всю эволюцию петалонам, найдется некоторое подтверждение "приземления" этих организмов. В течении некоторого времени эти стеблеобразные животные, крепившиеся ко дну с помощью грушевидных или дискообразных наростов у основания, в последствии отказались от данных приспособлений. Примером может служить фрактофус, чье тело полностью лежало на дне плашмя, осмотически поглощая питательные вещества, а возможно и сам бактериальный мат.
Конечно еще сложно говорить о фрактофусе, как о непосредственном предке проартикулят. Это было бы слишком смелое заявление. Однако некоторые предположения все-же имеются.
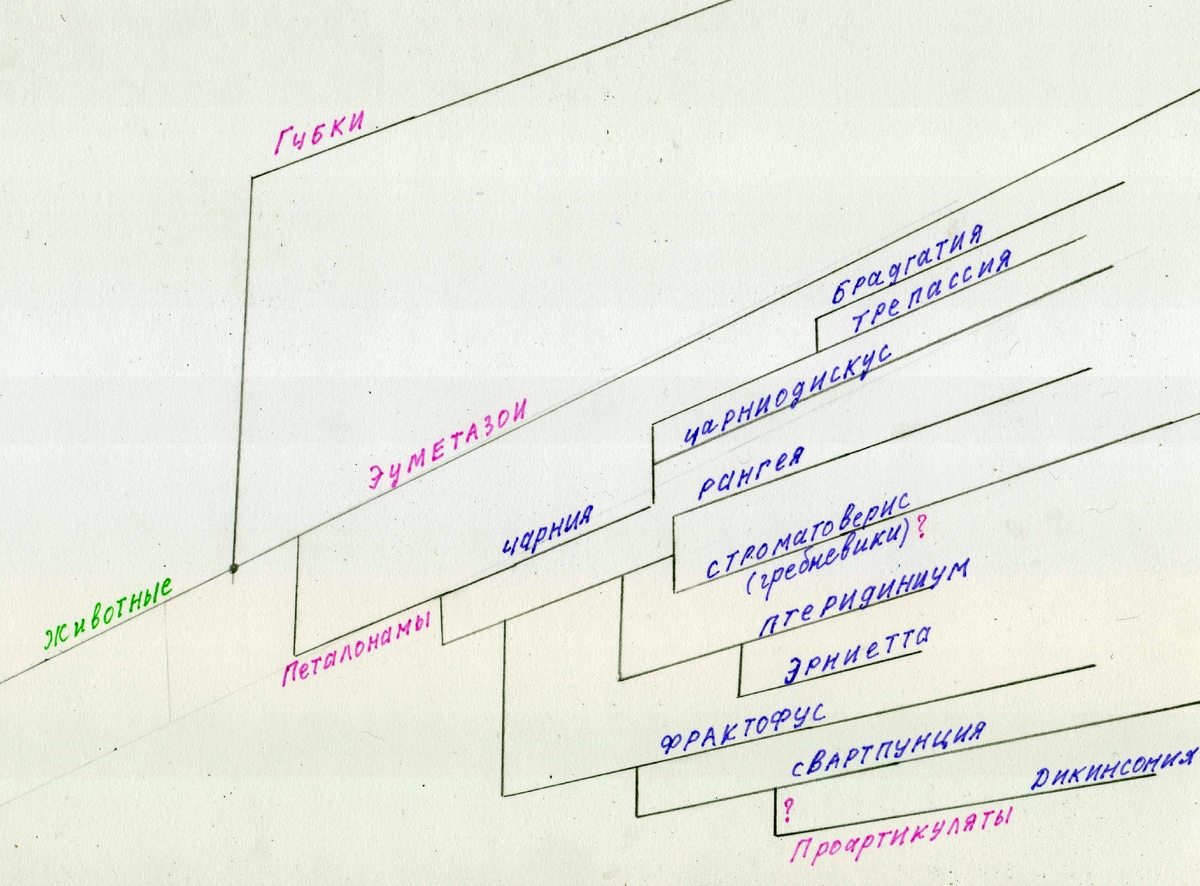
Конечно филогенетическое древо, изображенное сверху, не является полным и точным. Но вполне наглядно показывает предположительное родство первых проартикулят дикинсоний с типом петалонам. А что между ними общего?
Петалонамы, так же как и проартикуляты имеют скользящую двустороннюю симметрию. Только первые были неподвижными, а вторые все-таки как-то научились перемещаться. Вероятно, посредствам ресничного эпителия. Возможно со временем, у поздних проартикулят, развились иные способы перемещения. Об этом мы поговорим чуть позже. Но для чего петалонамам понадобилось научиться двигаться и стать в последствии типом проартикулят?
Движение вперед.
Ответ прост и сложен одновременно. Причины могли быть множественные. В фотических зонах было гораздо больше пищи, необходимого кислорода и согревающих солнечных лучей. Не исключено, что какие-то из петалонам могли освоить мелководья у прибрежных зон. Это могли быть какие-то лагуны, заливы или бухты. Там где вода стоит, где спокойно растет бактериальный мат - основные пастбища для первых многоклеточных животных. Но в эти лагуны надо было еще как-то попасть. Это весьма сложно сделать, когда ты на всю жизнь прикреплен к одному месту и лишь колыхаешься на волнах.
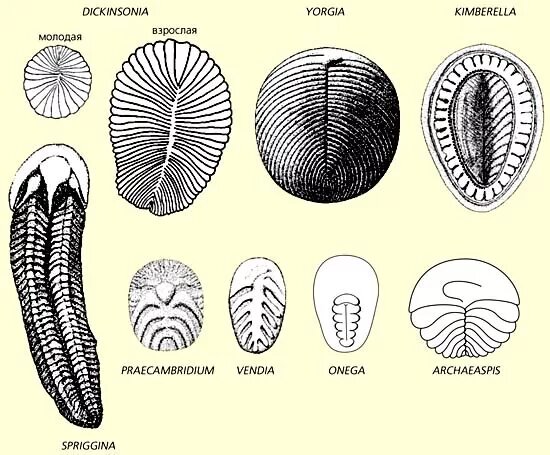
Не исключено, что ресничный эпителий имелся и у петалонам. Но использовался он только для ловли питательной взвеси в толще воды. Первые проартикуляты же научились с его помощью передвигаться, тем самым расширив функции этой группы клеток и своей ареал обитания.
Дальнейшая эволюция проартикулят вызывает не мало споров. Происхождение от них современны представителей сомнительно, но не исключено.
В ходе проведенных исследований:
1) Дана расширенная характеристика типа Proarticulata, изменен объем типа, который теперь включает 3 класса (1 новый), 18 валидных родов (8 новых) и 22 вида (9 новых).
2) Установлено, что проартикуляты были бентосными животными, населявшими органические субстраты, состоявшие из микробных матов и различных комплексов макроскопических эукариот. Места их обитания располагались в гидродинамически неспокойных обстановках в пределах фотической области от приливно-отливной зоны до глубин проникновения штормовых волн. Однако присутствие проартикулят в бентосных сообществах позднего эдиакария определялось распространением микробных матов, и мало зависело от глубины, вещественного состава минеральных грунтов, солености воды.
3) Впервые идентифицированы и описаны крайне своеобразные, не имеющие аналогов в палеоихнологии, следы питания проартикулят и следы двигательной активности, в том числе ползания, прикрепления к поверхности субстрата и бегства из-под внезапно выпавшего осадка. Определено, что в момент захоронения тела проартикулят были закреплены на субстрате.
4) Впервые описаны прижизненные повреждения, а также древнейшие в ископаемой летописи животных признаки регенерации части тела у дикинсоний. Реакция организма на нелетальные повреждения, протекание процесса регенерации и его отклонения подтверждают известное представление о локализации зоны роста на заднем конце тела.
5) Установлено, что рост дикинсоний был неопределенным и имел аллометрический характер. Выявленное сходство осуществления 41 процессов роста и регенерации у дикинсоний позволяет предположить, что оба процесса контролировались одной системой внутренних регуляторов, и что регенерация у дикинсоний является проявлением обычного роста этих организмов.
6) Выполнена новая реконструкция плана строения проартикулят (дикинсоний). Кроме ранее описанных признаков (выдержанная форма тела без боковых придатков и временных выростов, суббилатеральное поперечное расчленение, передне-задняя полярность), у этих организмов предполагается наличие дорсо-вентральной дифференциации, ресничного слизевыделяющего эпителия, подстилаемого базальной пластинкой, двух рядов слепых вентральных карманов, нервной системы диффузного типа, осевого опорного тяжа и мышц.
7) Выявленный набор признаков указывает на близость дикинсоний (и Proarticulata в целом) к Urbilateria – гипотетическому предку современных билатерально-симметричных животных. В то же время, присущий проартикулятам ансамбль элементов симметрии сближает их с Vendobionta
- группой своеобразных докембрийских радиат; а какие-либо ископаемые свидетельства преобразования изомерно построенных организмов в метамерные отсутствуют. Таким образом, отношение проартикулят к фанерозойским Bilateria пока остается неопределенным.
Не забывайте ставить лайк и подписывайтесь на канал. К вопросу об эволюции билатералий мы еще вернемся неоднократно. Оставляйте комментарий по теме. Всегда с вами канал "Страницы Земли".