Аннотация
Работа генома полностью описывается принципами облачного управления. Имеет устойчивость характерную для такой организации системы. Такой подход позволяет под другим углом зрения увидеть многие проблемы в медицине, в понимании старения.
Основная часть
Человечество создало «облако» из интернет сети. Появились программные продукты использующие вычислительные возможности множества участников и самих участников – как хранилища информации. Но природа и тут опередила нас на миллиарды лет. Геном – это облачно организованная и управляемая система. Нам известно, что ни один ген не отвечает за одно свойство или процесс организма, как верно и то - что любое свойство или процессинг в организме связан с работой множества генов. Рис 1. Это самая древняя организованная Облачная система. Образовалась она через самообучение и эволюцию своих инструментов – генов в системе генома. Изучая историю развития наших сетей мы, безусловно, увидим большое сходство с задачами и решениями которые уже были найдены миллиарды лет назад Живой материей.
Рис1
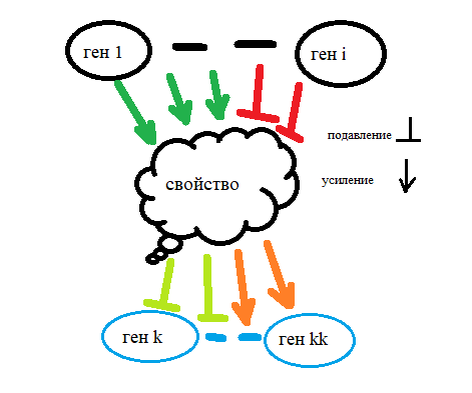
Мы, конечно же, не удивляемся уже разнообразию профилей экспрессии для разных фенотипов человеческого генома-организма. Не удивляемся и неожиданно большому генетическому разнообразию относительно «здоровых» мутаций в геномах человека. Не удивляемся неожиданно большой роли «мусора» (не транслируемых в белки последовательностей ДНК) в процессинге генома. В работе 3 впервые описывается процессорная модель организации работы генома. Т.е., можно рассматривать организацию процессинга геномов – как работу последовательной вычислительной машины. Нас удивляет только неожиданная устойчивость работы этого компьютера, который, ведет управление в достаточно жестких условиях работы, в реальном времени. При этом, десятки тысяч процессов в клетке идут параллельно, находясь в сильной или слабой связи между собой и с внешними событиями. При всем при этом, в клетке есть внутренние водители множества ритмов. Теперь, мы наблюдаем не совсем обычное поведение конкретного сообщества клеток и даже отдельных из них. Так, мы отмечаем существенные отличия на фоне «стандартных» свойств во времени: 1- пролиферации клеток или 2 – экспрессии генов. Необычные свойства могут лежать и в области их свойств изменчивости и миграции. Хотя, всё это прямо следует из обнаруженного разнообразия фенотипов.
Отличие работы этого процессора от привычных нам архитектур наших процессоров связано с историей - эволюцией развития живой материи. Там, реальное время было залогом эффективности, а не точность и даже достаточность. Параллельность процессов компенсировало низкую скорость единичных шагов процессора. При этом основная часть работы «софтвера» ведется в очень медленных – биохимических процессах. И, естественно, что не они отвечают за работу в реальном времени. Они просто не имеют достаточной скорости и «инструментария» для исполнения, грубо говоря, исполнительные устройства - не электронные быстродействующие компоненты и механизмы, а сами биохимические процессы. Таким образом, эволюция развила длинную цепочку от сигнала раздражителей или водителей до реагирования организма, движения мышц и т.п.. И всё же, тут речь, о метаболистической части управления в клетках. Метаболизм жестко завязан на уже легко наблюдаемые макроявления: пролиферации клеток, органоидов, экспрессии генов и т.п. .
Нужно разделять, как разделены во времени и пространству ядерные ДНК-РНК – процессинги и клеточные РНК-белковые и клеточные ко-процессинги. Именно в силу относительной замедленности связи ко-процессинга с ДНК-процессингом, развилась внутренняя «быстрая» сеть реагирования в реальном времени. Она образована медиаторно-белковой-РНК системой переключателей и путей с обратной связью, которая призвана, не дожидаясь полной отработки «командного» центра ДНК-компьютера, отработать возникающие внутренние диссонансы с внешними стимулами. Это позволяет клетке быстро реагировать на важные, с точки зрения выживаемости и функционала изменения – диссонансы, во внешних сигналах (любого типа) уже на этапе прохождения по сигнальным тропам от рецепторов внешней мембраны к ядру. С последующей корректировкой – поправкой, которая будет уже результатом интегративной работы ДНК-процессинга, принимающего во внимание все обстоятельства - предсостояние, состояние эпигенетической и фенотипической настройки, управляющего фона водителей ритмов и т.п. .
При исследовании различных реакций различных клеток на внешние воздействия, исследователи наблюдают результаты в виде сложных временных профилей экспрессии генов и продуктов процессингов и метаболизма. Только в последние время появились работы посвященные глубоким исследованиям экспрессии многих участников процессингов в единичных клетках. Серьезным ограничением в достоверности является вынужденная изоляция исследуемых клеток от естественной среды, которая преодолевается простым увеличением числа и времени экспериментов с её вариантами. Поэтому, планирование экспериментов – это серьезнейшая задача, она содержит в себе потенциал во множество порядков сокращение времени и информативности экспериментов. И тут простым перебором вариантов дело не решается, сотни тысяч «участников» процессинга, влияние временных факторов, предсостояния и водителей ритмов – приводят исследователей к мысли о сжатии исследования, за счет его информативности. Невообразимая вариативность требует теоретических основ для своего эффективного изучения.
При том, что генетические исследования это вызов будущему развитию Больших Данных и методам их обработки, тут присутствует большой компонент необходимости установления достоверности и достаточности получаемой информации. Пересмотр парадигмы управления – путь, который, наверное, уже чувствуется многими большими исследователями в генетике. К этому нас приводят и результаты выше упомянутой работы (1). Особенно неожиданно участие в достаточно медицинских -разнородных патологиях одних и тех же важных объектов (микроРНК, генов, белков и других молекулярных инструментов процессингов). Но теоретиков в области Генетики – нет…
Тут, мы подходим к первому камню на пути понимания работы более 25 тыс транслируемых генов и всего, такого нужного - «мусора» в геноме клетки. Уже упомянутое выше, ставшим первым поразительным открытием широкого секвенгирования генома человека: разнообразие в фенотипах человека. Естественным продолжением стало: открытие большого фенотипического разнообразия в профилях экспрессии генов и микроРНК клеток одних и тех же органах. Все исследования, до сего момента, базировались на презумпции существования единого «идеального» стандарта. Оно соответствовало понятию – совершенно здорового организма. Эти открытия просто перевернули наши представления об этих стандартах. Их просто не оказалось. В чем дело ?! А дело в том, что геном сложных организмов, в процессе эволюции, развил большую сеть скомпенсированных воздействий на главные циклы метаболизма организма и его клеток. Пониманию этого явления способствовало развитие Информационных Технологий (ИТ) и, таким образом - родился используемый мной термин «облачное» управление (ОУ), в применении к генетическому процессингу в клетках. Этот термин зеркально отражает его смысл в ИТ. Получается так, что в процессинге клетки ЕСТЬ ключевые константы, которые ПРЕСЛЕДУЮТСЯ этими процессингами – как действительно: ОПРЕДЕЛЯЮЩИЕ. Вообще говоря, это естественно следует из работы 1. Ведь «компьютерная» организация управления предполагает наличие процессинговых «водителей». Другое дело, где их искать ? Эта, казалась бы, чисто теоретическая задача, находит свое прямое применение в некоторых упорных патологиях: фиброзах, перерождении тканей, онкологии, воспалениях, старении организма и др. .
Т.о., тут необходимость теории совместила как задачи изучения процессинга, так и задачи лечения (эффективного управления) патологий. От этого задачи, по своей сути, не изменились, но, включили в себя и «больные» варианты, умножив тем самым трудности изучения. С другой стороны, патологии косвенно указывают на инструменты их водителей.
Рассматривая вопрос о развитии лекарственной устойчивости в онкологии и других патологиях, очевидно, что рассмотрение её с точки зрения микроэволюции – абсурдно. Громкие обобщения делаются из генетических исследований онкологии. А исследования эти - достаточно обширны. Как уже писалось выше, что относиться к управлению (процессингу) генома клетки НУЖНО - как к облачной системе. Т.е.: ни один ген не обеспечивает одно свойство клетки или организма и ни одно явление и свойство в организме и клетке - не связано с одним геном. Более того, даже во времени, здоровый организм - существует в "облаке" возможностей и компенсаторных генетических троп, предлагая организму (клетке) разнообразные варианты реагирования. Кстати, некоторые из них заканчиваются патологиями и онкологией. Можно констатировать, что наши знания о функционировании троп генов здорового организма или, по крайней мере, не онкогенных образований, крайне недостаточны, но и они не дают основания для применения эволюционного аппарата для предсказания их развития.
Что уж говорить об онкологии - резко отличающимся состоянии своих же клеток - под общим названием: ОНКОЛОГИЯ. С одной стороны: клетки ведут себя как быстро регенерирующие или эмбриональные ткани, с другой, этот процесс не имеет границ, в отличие от нормальных. Сравнивать происходящие явления с эволюцией можно только на основании расходящихся линий гетеронаселений у одних фенотипов. А мы наблюдаем жесткую приверженность одних и тех же фенотипов онкологии к предопределенным трансформациям первичных онкообразований. И это позволяет последовательно искать и часто находить новые ключевые точки сопротивления лекарственным воздействиям. Это совершенно не похоже на свободное течение эволюции, а скорее, на движение в канале облачных возможностей данного фенотипического типа определенных видов онкологий. Что и служит основой для общего и продуктивного подхода к поиску новых дополнительных лекарственных средств.
То - что генетическая нестабильность имеет место - абсолютно не означает произвольного течения мутационных изменений в онкоклетках. Нестабильность генома- просто осложняющий фактор, он поддается определению, а не "эволюционный" вариант, который по условию, ничего не привносит в планирование работы по этому направлению. Наоборот, вносит презумпцию произвольной трансформации. Что глубоко различается с существом текущих процессов.
Во-первых: потому что наблюдаемый «тренд» мутаций - практически одинаков во всех случаях, он обеспечивается типом онкоклеток, микроокружением и фенотипом (сопутствующими мутациями).
Во- вторых: гетеронаселение конкретной популяции представлено не разнообразными «эволюционными наборами» КК, а скорее линиями, похожими на линии трансформации стволовых клеток с искаженными мутациями и представленными как дифференцированными (ДК) так и стволовоподобными онкоклетками (КСК).
В- третьих: Нет статистического «набора» однотипных онкоклеток (КК), и не наблюдается набор «эволюционного опыта» в виде таких клеток. Они скорее – исключение, чем пример.
В – четвертых: в самой популяции конкретного онкообразования в самом начале обработки уже находят эти устойчивые к лекарствам клетки .
На этом фоне появляется вопрос: что же движет развитием лекарственной устойчивости? Как показал опыт, попытка «перекрытия» уж жизненно-важных путей выживания КК и КСК – через инструменты процессингов, приносят временный и, очевидно, не полный успех. Уже в самой онкопопуляции существуют устойчивые к лекарственной обработке КСК. С одной стороны: идет простое увеличение их числа. С другой – именно ввиду их стволовых свойств они дают свои «деревья» - линии КК, многие (!) из которых изначально устойчивы к используемому лекарственному воздействию. Поэтому, часто, совмещение нескольких лекарств дает синергетический или аддитивный эффект. Чтобы ответить на этот вопрос, вернемся к способу управления процессингом клетки, а он - облачный. Значит, устанавливая препятствия на отдельных путях метаболизма КК, мы просто заставляем облачное управление(ОУ) использовать компенсаторные пути обхода препятствия. Другое дело, что при этом, КК развиваются по не предусмотренному НАМИ сценарию. Но этот сценарий ЕСТЬ в списке вариантов стволовых клеток (СК), а значит и КСК. По-крайней мере: иммунная система его пропускает, механизмы апоптоза и другие защитные механизмы – не срабатывают. Мы находим молекулярные и генетические пути, обеспечивающие эти явления. Весь вопрос в том – что может выдержать организм и клетки-соседи КК, многие из которых сами используют такие механизмы защиты. При всем при этом, мы видим, что КК сильно отличаются по своим функциональным свойствам от нормальных СК и ДК (но пользуются они тем же ДНК!). Конечно, ясно, что у организма нет «супервайзера» функционалов клетки, но есть супервайзеры свойств клетки. Часть из них это: системы BCL, BCL-x, SNAIL, PTEN, p53- p21, Vav, BRCA-1,2… тысячи микроРНК, сотни лонгРНК и т.п. . Список впечатляющий и, как показывает история, почти бесконечный.
На этом пути «стандартные» точки – миф, но процессинговые константы – константы свойств, реально существуют и мы, косвенно, ориентируемся на них в наших субъективно-объективных оценках результатов экспериментов над патологическими клетками. Отсроченные результаты работы этих истинных водителей свойств клеток и есть наша цель.
Литература:
1. “Adaptation and selection shape clonal evolution of tumors during residual disease and recurrence” Andrea Walens, Jiaxing Lin, et all, Nature Communications volume 11, Article number: 5017 (2020)
2. “Cell plasticity and senomic instability in cancer evolution” Yan Xu, Hui Li, Fan Yang, et all Genome Instability & Disease volume 1, pages301–309(2020)
3. «Evolution origin of modern genome» Astashkin Sergey N 2005г http://www.science-journals.eu/