Введение
Человеческая жизнь начинается с события, одновременно микроскопического и грандиозного, в результате которого две специализированные клетки сливаются и дают старт программе, длящейся примерно тридцать восемь недель. За этот срок из единственной зиготы возникают триллионы клеток, организованных в ткани, органы и системы, способные поддерживать дыхание, питание, движение и мышление. Ещё полвека назад эмбриональное развитие описывали как последовательность морфологических перестроек, опирающуюся главным образом на световую микроскопию и сравнительную анатомию. Сегодня благодаря молекулярной биологии, геномике, передовым методам визуализации и культивирования эмбрионов мы расшифровываем инструкции, записанные не только в последовательности ДНК, но и в её эпигенетических метках, пространственных градиентах сигнальных молекул и даже механических напряжениях тканей.
Современная эмбриология интегрирует данные секвенирования РНК одиночных клеток, редактирования генома, органоидных технологий и неинвазивных пренатальных тестов, чтобы понять, каким образом достигается поразительная точность и пластичность развития. Параллельно общество сталкивается с глубокими этическими вопросами, порождёнными появлением искусственных гамет, систем внеутробного вынашивания и возможностью редактировать геном зародышевой линии. В этой статье мы проследим путешествие от оплодотворения до рождения, останавливаясь на самых захватывающих достижениях последнего десятилетия и критически переосмысляя устаревшие метафоры, которые до сих пор встречаются в популярных изложениях. Задача состоит не только в том, чтобы удивиться сложности человеческого зачатка, но и в том, чтобы понять, как знания о внутриутробном программировании здоровья могут изменить медицинскую практику и наше отношение к самым первым этапам существования.
Глава 1. Встреча двух геномов: оплодотворение и активация зиготы
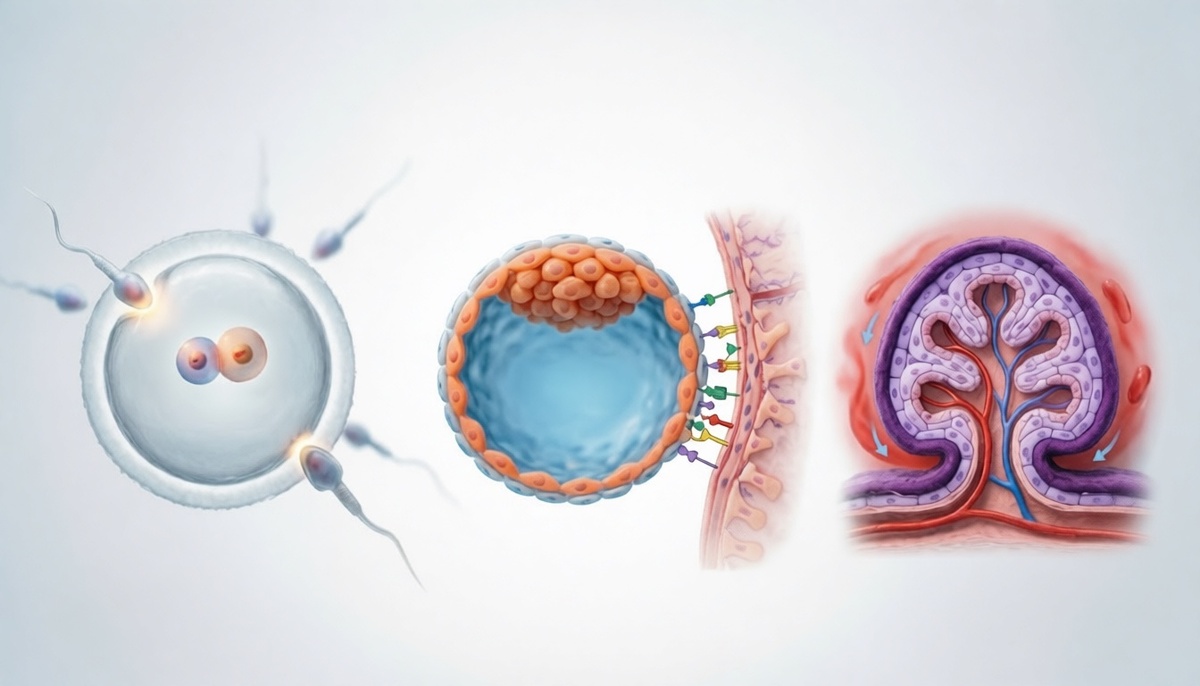
Оплодотворение традиционно представляют как гонку сперматозоидов, однако на клеточном уровне это тщательно синхронизированный диалог между мужской и женской гаметами. После проникновения сперматозоида через прозрачную оболочку яйцеклетки в цитоплазме ооцита распространяется волна ионов кальция, которая блокирует полиспермию и запускает завершение второго мейотического деления. В результате формируются два пронуклеуса — материнский и отцовский, которые мигрируют навстречу друг другу, обмениваются регуляторными сигналами и затем синхронно разрушают ядерные оболочки, смешивая хромосомы. Этот момент, называемый сингамией, знаменует появление зиготы с уникальным диплоидным набором из сорока шести хромосом.
Новые исследования показали, что родительские геномы привносят не только разные аллели генов, но и неодинаковые эпигенетические репертуары. Отцовские хромосомы приходят компактно упакованными с помощью протаминов и после оплодотворения активно деметилируются, тогда как материнские сохраняют значительную часть своих меток до начала дробления. Явление геномного импринтинга, при котором экспрессия гена определяется его родительским происхождением, объясняет, почему наличие обоих геномов необходимо для нормального развития. Нарушения импринтинга, возникающие при неправильной эпигенетической модификации локусов, лежат в основе синдромов Прадера–Вилли, Ангельмана и некоторых форм задержки внутриутробного роста.
В первые дни деления, именуемые дроблением, бластомеры становятся всё меньше, а общий объём зародыша почти не меняется. У человека в это время реализуется особый тип регуляции, при котором собственный геном эмбриона транскрибируется лишь частично, а основную нагрузку несут материнские матричные РНК, запасённые в ооците. Переход от материнского к зиготному контролю развития происходит на стадии четырёх–восьми клеток и считается одним из самых уязвимых этапов; сбои в этой смене управления являются ведущей причиной остановки развития до имплантации. Углублённое изучение транскриптомов единичных бластомеров позволило выделить молекулярные маркёры, которые помогают эмбриологам отбирать наиболее перспективные эмбрионы в программах ЭКО.
К концу третьих суток зародыш представляет собой морулу — плотный шар из восьми–шестнадцати компактизированных клеток. Компактизация усиливает межклеточные контакты и создаёт первую в жизни человека функциональную асимметрию: внешние клетки приобретают свойства трофэктодермы, отключая часть генов плюрипотентности и активируя программу будущего плацентарного роста. Этот первичный выбор судьбы, происходящий ещё до имплантации, подчёркивает, что дифференцировка начинается гораздо раньше, чем считалось десятилетия назад, и опирается на комбинаторное действие транскрипционных факторов, таких как CDX2 для трофэктодермы и OCT4 для внутренней клеточной массы.
Глава 2. Бластоциста и молекулярный диалог, разрешающий имплантацию
Примерно на четвёртый–пятый день после оплодотворения внутри морулы начинает накапливаться жидкость, и зародыш преобразуется в бластоцисту — полую сферу, стенка которой образована трофэктодермой, а на одном из полюсов к ней примыкает плотный узелок внутренней клеточной массы. Жидкость создаёт бластоцель, и давление внутри полости в сочетании с ферментативным размягчением оболочки приводит к высвобождению бластоцисты из прозрачной зоны — процессу, называемому хэтчингом. При нарушении хэтчинга эмбрион не способен вступить в контакт с эндометрием, что делает его нежизнеспособным; именно поэтому в клиниках ЭКО иногда применяют лазерный вспомогательный хэтчинг.
Имплантация начинается на шестые–седьмые сутки и представляет собой не механическое врастание, а сложный молекулярный диалог между клетками трофэктодермы и выстилкой матки, подготовленной под действием прогестерона. Трофобласт экспрессирует молекулы адгезии, L‑селектин и интегрины, а также выделяет факторы роста и цитокины, перестраивающие строму эндометрия и привлекающие иммунные клетки матери, которые парадоксальным образом способствуют закреплению зародыша. Синцитиотрофобласт — продукт слияния клеток трофэктодермы — внедряется в толщу слизистой, формируя лакуны, которые вскоре заполнятся материнской кровью.
Современные исследования раскрывают имплантацию как уникальный иммунологический компромисс. Клетки трофобласта практически не экспрессируют классические молекулы главного комплекса гистосовместимости класса I, вместо этого они несут неклассические молекулы HLA‑G и HLA‑E, которые взаимодействуют с рецепторами естественных киллеров и макрофагов матки. Эти взаимодействия не только предотвращают отторжение генетически наполовину чужеродного эмбриона, но и направляют ремоделирование спиральных артерий матки, необходимое для глубокой плацентации. Сбой в этом диалоге считают одним из патогенетических узлов преэклампсии и привычного невынашивания беременности.
С развитием технологий культивирования эмбрионов до стадии поздней бластоцисты и появлением методов секвенирования открытого хроматина отдельных клеток учёные смогли составить атлас транскриптомов и эпигенетических ландшафтов для каждой линии: трофэктодермы, эпибласта и примитивной энтодермы. Оказалось, что уже в момент имплантации в клетках внутренней массы начинают экспрессироваться гены, регулирующие будущую ось тела, и даже выявляются предшественники внеэмбриональной мезодермы. Эти данные заставляют пересмотреть представление о том, что судьба клеток окончательно фиксируется только после гаструляции.
Глава 3. Гемоплацентарный барьер и уникальная эндокринная фабрика
Формирование плаценты начинается одновременно с имплантацией и выходит на плато к концу первого триместра, однако её структура продолжает тонко настраиваться до самых родов. Ворсины хориона, покрытые двухслойным эпителием — цитотрофобластом и синцитиотрофобластом, — врастают в материнскую децидуальную оболочку, ветвятся и омываются материнской кровью в межворсинчатом пространстве. Гемоплацентарный барьер на начальных этапах содержит несколько слоёв: синцитий, цитотрофобласт, его базальную мембрану, соединительную ткань ворсины и эндотелий плодового капилляра. К концу беременности цитотрофобласт регрессирует, что ускоряет обмен газами и питательными веществами, но одновременно делает плаценту более уязвимой для некоторых вирусов, таких как цитомегаловирус и вирус Зика.
Сосуды пуповины, заключённые в вартонов студень, транспортируют кровь в двух направлениях. Единственная пупочная вена доставляет к плоду кровь, обогащённую кислородом, от плаценты, а две пупочные артерии возвращают дезоксигенированную кровь обратно для очистки и насыщения. Ультразвуковая допплерометрия этих сосудов и маточных артерий стала рутинным инструментом оценки гемодинамики плода и позволяет рано предсказывать задержку роста и гипоксию. Отклонения в резистивных индексах часто опережают клинические симптомы и дают возможность своевременно изменить тактику ведения беременности.
Плацента секретирует целый спектр гормонов, без которых невозможна адаптация материнского организма и рост плода. Хорионический гонадотропин, вырабатываемый в первые недели, поддерживает жёлтое тело; плацентарный лактоген модулирует метаболизм глюкозы и липидов; прогестерон и эстрогены обеспечивают гемодинамические изменения и подготавливают молочные железы. Современные протеомные и липидомные анализы выявили, что трофобласт продуцирует также экзосомы, нагруженные микроРНК и сигнальными белками, которые попадают в материнскую циркуляцию и, вероятно, участвуют в системной иммунной модуляции.
Эволюционное значение плаценты выходит далеко за рамки простого физического контакта матери и плода. Сравнительная геномика показала, что многие гены, экспрессирующиеся в трофобласте, произошли от эндогенных ретровирусов и были кооптированы для управления слиянием клеток в синцитий и обеспечения толерантности. Уникальные молекулярные особенности плаценты делают её не только самым быстро стареющим органом человека, но и важнейшим объектом для изучения механизмов старения и регенерации. Понимание того, почему плацента избегает злокачественной трансформации гораздо реже других тканей, может открыть подходы к терапии опухолей.