
Древние пшеницы лучше усваиваются организмом чувствительным к глютену.
В данной статье опубликованной в журнале Foods в 2020м году ученые показывают, что однозернянка более безопасный продукт даже для людей страдающих целиакией.
Аннотация: Целиакия (CeD) - аутоиммунная энтеропатия, вызываемая иммуногенными пептидами глютена, которые высвобождаются в процессе переваривания пшеницы в желудочно-кишечном тракте. Нашей целью было идентифицировать пептиды, содержащие Т-клеточные эпитопы, после переваривания ex vivo древней (однозернянка, спельта и эммер) и обычной (гексаплоидной) пшеницы (Fram, Bastian, Børsum и Mirakel) с использованием желудочно-кишечных соков человека. Пшеничную кашу переваривали с использованием статической модели ex vivo. Пептиды, выделившиеся после 240-минутного переваривания, анализировали с помощью жидкостной хроматографии в сочетании с масс-спектрометрией высокого разрешения (HPLC-ESI MS/MS). При переваривании Ex vivo у древних сортов пшеницы (однозернянка (n = 38), спельта (n = 45) и эммер (n = 68)) высвобождалось меньше пептидов, содержащих Т-клеточные эпитопы, по сравнению с обычными сортами пшеницы (Fram (n = 72), Børsum (n = 99), Bastian (n = 155) и Mirakel (n = 144)). Ни в одном из переваренных сортов пшеницы не было обнаружено ни иммунодоминантных 33-мерных и 25-мерных α-глиадиновых пептидов, ни 26-мерного γ-глиадинового пептида. Таким образом, пищеварительный сок человека был способен переваривать 33-мерный и 25-мерный α-глиадин, а также 26-мерные пептиды, полученные из γ-глиадина, в то время как их фрагменты все еще содержали наивные Т-клеточно-реактивные эпитопы. Хотя древняя пшеница выделяла меньше иммуногенных пептидов после переваривания человеком ex vivo, они по-прежнему высокотоксичны для пациентов с целиакией. Более широкое использование этих древних сортов пшеницы может, тем не менее, снизить частоту возникновения целиакии CeD.
Ключевые слова: целиакия; пшеница; переваривание ex vivo; Т-клеточный эпитоп; иммуногенный пептид
Введение
Белки пшеницы вызывают реакции гиперчувствительности (аллергия/непереносимость). Целиакия (CeD) - аутоиммунная реакция гиперчувствительности, вызываемая глиадинами пшеницы у генетически предрасположенных людей. Популяционный скрининг показал, что распространенность CeD составляет 1-2% в Европе и США [1,2], но многие пациенты еще не диагностированы [3]. Помимо пшеничного глютена (глиадин и глютенин), пациенты с CeD реагируют на структурно родственные белки во ржи (секалины), ячмене (гордеины) и редко в овсе (авенины) [4]. Вызванное глютеном воспаление кишечника ведет к гиперплазии клеток крипт тонкой кишки, атрофии ворсинок и нарушению всасывания питательных веществ [5]. Доминирующая генетическая предрасположенность к CeD - наличие аллелей человеческого лейкоцитарного антигена (HLA) DQ-2.5 или DQ8 [6]. В мире около 90% пациентов с CeD экспрессируют гетеродимер HLA-DQ 2.5 (состоящий из HLA-DQA*0501 и HLA-DQB*0201), остальные экспрессируют HLA-DQ8, и лишь у нескольких экспрессируется 1 из цепей HLA-DQ-2.5 либо в HLA-DQ2.2, либо в -DQ7.5 alleles [7]. Хотя HLA - наиболее важный и необходимый генетический фактор риска, более 39 других локусов связаны с развитием CeD [8].
Молекулы HLA-DQ, ассоциированные с CeD, связывают пептиды глиадина, которые были дезаминированы тканевой аутоантигенной трансглутаминазой-2 (tTG2) с высокой аффинностью. Однако выбор Т-клеточных эпитопов, реагирующих с глютеном, зависит, по меньшей мере, от трех факторов: (1) устойчивости к протеолитическому расщеплению, (2) специфичности к дезаминированию tTG2 и (3) связывающих свойств с HLA-DQ-2.5/8.
Основными белковыми группами в пшенице являются запасающие белки глиадин и глютенин, составляющие 80% от количества белка в зерне обычной пшеницы [9]. Они богаты глютамином и пролином, что делает глютен очень устойчивым к протеолитической деградации в ЖКТ, в результате чего образуются длинные пептиды длиной от 15 до 50 остатков [10]. Пример: 33-мерный фрагмент α-глиадина (LQLQPFPQPQLPYPQPLPYPQPQLPYPQPQPF) который считается 1м из главных «виновников» CeD [10,11] поскольку он содержит 6 перекрывающихся Т-клеточных эпитопов. Но было идентифицировано несколько других длинных пептидов, содержащих CeD-ассоциированные Т-клеточные эпитопы [12–14].
Обыкновенная пшеница (Triticum aestivum) развилась в результате гибридизации между тетраплоидным видом T. turgidum (AABB) и диплоидным видом Aegilops tauschii (DD) [15]. Обыкновенная гексаплоидная пшеница (AABBDD) хорошо адаптируется, растет более устойчиво и имеет благоприятный состав глютена для промышленного производства по сравнению с предшественницами [16]. Но разница в составе глютена между диплоидными (AA), тетраплоидными (AABB) и гексаплоидными (AABBDD) сортами может влиять на расщепление при пищеварении [17]. Замена обычной пшеницы на однозернянку (Triticum monococcum) отсрочит начало CeD у предрасположенных к HLA-DQ2.5/-8 родственников 1й степени, т.к. содержит меньше предполагаемых Т-клеточных эпитопов [14] (т.к. у диплоидной и тетраплоидной пшеницы нет генома Dс 33-мерным фрагментом α-глиадина).
Цель исследования - сравнить пептидные профили и идентифицировать глютеновые пептиды, полученные ex vivo из 7 различных сортов пшеницы с использованием желудочно-кишечных соков человека. Для обработки использовали древнюю пшеницу (однозернянка, спельта и эммер) и сорта обычной пшеницы, выращиваемые в Норвегии в 1900-1930 г. (Børsum, Fram) и в 1990-2020 г. (Bastian, Mirakel). Поскольку активность и специфичность пищеварительных ферментов, в т.ч. трипсин, различны в разных источниках [18], использование пищеварительных ферментов человека более точно отразит ситуацию in vivo по сравнению с ферментами бычьего или свиного происхождения, которые применялись в большинстве опубликованных на сегодняшний день исследований in vitro.
Пептиды, высвобождающиеся при расщеплении, были идентифицированы с помощью высокоэффективной жидкостной хроматографии/ионизации электрораспылением высокого разрешения/тандемной масс-спектрометрии (HPLC-ESI-MS/MS). Полученные пептиды сравнивали с 9ю аминокислотными участками ядра, распознаваемыми CeD-ассоциированными CD4+ Т-клетками [19]. Кроме того, был проведен мониторинг 33-мерного и 25-мерного α-глиадинов и 26-мерного γ-глиадинового иммунодоминантного пептида, поскольку они состоят из нескольких перекрывающихся Т-клеточно-реактивных эпитопов.
Материалы и методы
Отбор проб пшеницы
Пшеница была собрана в 2017 г. с экспериментального поля (Vollebekk Research Farm, Norwegian University of Life Sciences, Ås, Norway). Все виды и сорта пшеницы выращивались на одном и том же поле на участках площадью 4.5 м2. По достижении зрелости с каждого участка было собрано примерно по 50 початков каждого образца. Были отобраны образцы пшеницы всех яровых сортов, включая виды древней пшеницы: однозернянка (диплоидная, AA), эммер (тетраплоидная, AABB) и спельта (гексаплоидная, AABBDD), а также 4 отобранных сорта обычной пшеницы (гексаплоидная, AABBDD). Эти 7 различных сортов пшеницы (Таблица 1) в дальнейшем называются “типами пшеницы”. Перед обмолотом и очисткой собранную пшеницу выдерживали при влажности ниже 15% при температуре 30°C в течение 3 дней (Perten Instruments AB, Hägersten, Sweden).
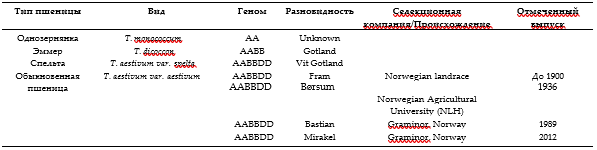
Характеристика пшеницы
После обмолота зерна однозернянки, эммер и спельты были очищены вручную. Перед дальнейшим анализом образцы были измельчены до состояния цельнозерновой муки на аппарате Falling Number Laboratory 3100 с ситом толщиной 0,8 мм (Perten Instruments AB, Hägersten, Sweden). Размер ядер измеряли по весу на тысячу ядер (TKW). Зерна подсчитывали с помощью счетчика семян Elmor C1 (Elmor Ltd., Schwyz, Switzerland), и представляли в виде веса в граммах на тысячу зерен. Содержание влаги в зерне определяли путем сушки зерен в течение 24 часов при температуре 105◦C. Кроме того, содержание азота в образцах пшеницы измеряли с помощью микро-метода Кьельдаля (Kjeltec™ 8400, Tecator, Foss, Hillerød, Denmark), а содержание белка в пшенице определяли с использованием коэффициента Кьельдаля 5,7. Общее содержание крахмала анализировали с помощью набора Megazyme (K-TSTA-100A 08/19, Megazyme, Bray, Ireland) [20]. Кашу готовили путем смешивания цельнозерновой муки и воды (1:20 вес/объем), затем нагревали при 100 ◦C в течение 10-15 мин, гомогенизировали, охлаждали и хранили при 4◦C до переваривания ex vivo
Переваривание пшеничной каши Ex vivo
Желудочный и дуоденальный соки человека собирали в соответствии с Ulleberg и соавт. [21] путем аспирации у здоровых добровольцев (n = 20) в Lovisenberg Diaconal Hospital, Norway. Все испытуемые сообщили об отсутствии симптомов CED и дали свое информированное согласие на включение в исследование до его начала. Это стремление было одобрено Regional Committees for Medical and Health Research Ethics (REK 2012/2230 и 2012/2210) в Норвегии. Вкратце, гибкая силиконовая трубка с тремя просветами была введена через нос или рот в антральный отдел желудка и двенадцатиперстную кишку под руководством гастроскописта. Одновременно с аспирацией желудочной и дуоденальной жидкостей непрерывно вводили изотонический стимулирующий раствор (17,5 г/л сахарозы, 450 мг/л NaCl, 800 мг/л L-фенилаланина и 575 мг/л L-валина в воде) (100 мл/ч). Аспираты объединяли и хранили при температуре −20 ◦C, а затем при −80 ◦C [21]. Ферментативную активность пепсина и трипсина определяли в соответствии с исследованиями Minekus и соавт. [22]. Переваривание с помощью ферментов ЖКТ человека проводилось в соответствии со стандартизированной консенсусной моделью [22] с некоторыми изменениями. Аликвоту овсяной каши (1 г с содержанием белка приблизительно 5 мг/мл) смешивали в соотношении 1:1 (вес/объем) со слюнной жидкостью (SSF), содержащей α-амилазу (75 U/mL, Sigma Aldrich) и инкубировали в течение 2 мин, имитируя пероральную фазу. Фазу переваривания в желудке проводили путем добавления имитационной желудочной жидкости (SGF) с желудочным соком человека (HGJ) (активность пепсина 2000 Ед/мл) к пероральной фазе (1:1, по объему) и доведения рН до 3,0 путем добавления 1 М HCl. Образцы инкубировали на водяной бане при температуре 37 ◦C при мягком магнитном перемешивании в течение 120 минут. Фазу дуоденального переваривания проводили путем добавления имитационной кишечной жидкости (SIF) содержащей дуоденальный сок человека (HDJ) (активность трипсина 100 ед/мл) к образцу из желудка (1:1 по объему). рН доводили до 7,0 добавлением 1 М NaOH и образцы инкубировали на водяной бане при температуре 37◦C еще 120 мин при магнитном перемешивании, затем добавляли 5 mM Pefabloc® (Sigma Aldrich, St. Louis, MO, US). Переваривание проводили параллельно, и все образцы сразу же хранили при температуре −20 ◦C до дальнейшего анализа.
Пептидный профиль с помощью HPLC-ESI MS/MS
Перед проведением MS анализа HPLC-ESI MS, осадки (100 мкл) были опреснены с помощью колонки C18 spin (Thermo Scientific, San Jose, CA, USA), в соответствии с инструкциями производителя, элюируя 70% ацетонитрилом (по объему)/0.1% трифторуксусной кислотой (TFA). MS-анализ проводили с использованием масс-спектрометра Q Exactive Orbitrap (Thermo Scientific, San Jose, CA, USA), в режиме онлайн в сочетании со сверхвысокопроизводительным прибором жидкостной хроматографии Ultimate 3000 (Thermo Scientific, San Jose, CA, USA). Очищенные пептиды разводили в 50 мкл 0,1%-ного (по объему) раствора муравьиной кислоты, пропускали через предварительную колонку длиной 5 мм и внутренним диаметром 300 мм (LC Packings, San Jose, CA, USA) и разделяли с помощью колонки EASY-Spray™ PepMap C18 (2 мкм, 15 см–75 мкм; частицы размером 3 мм; размер пор 100 Å (Thermo Scientific, San Jose, CA, USA)). Элюент А состоял из 0,1% муравьиной кислоты (по объему) в воде Milli-Q, а элюент В состоял из 0,1% муравьиной кислоты (по объему) в ацетонитриле. Колонку уравновешивали 5%-ным элюентом B. Пептиды разделяли с помощью градиента элюента В 4-40% в течение 60 мин (300 нЛ/мин). Масс-спектрометр работал в режиме, зависящем от данных, и все спектры MS1 были получены в режиме положительной ионизации путем сканирования в диапазоне 1800-350 м/с. Максимум 10 наиболее интенсивных ионов MS1 были фрагментированы в режиме MS/MS. Разрешающая способность была установлена на уровне 70 000 по всей ширине при половинном максимуме (FWHM), используя мишень автоматической регулировки усиления (AGC), равную 1 × 106 ионам, и 100 мс в качестве максимального времени инжекции ионов (IT) для генерации спектров предшественников. Спектры фрагментации MS/MS были получены с разрешающей способностью 17,500 FWHM и для предотвращения повторной фрагментации наиболее распространенных ионов использовалось динамическое исключение в течение 10 секунд. Ионы с одним или более чем шестью зарядами были исключены из процесса фрагментации. Спектры были получены с использованием программного обеспечения Xcalibur Software 3.1 version (Thermo Scientific, San Jose, CA, USA).
Идентификация спектров МС-анализа
Пептиды были идентифицированы по спектрам MS/MS с использованием Proteome Discoverer 2.1 software (Thermo Scientific, San Jose, CA, USA), основанного на алгоритме Sequest searching. Поиск был таксономически ограничен базой данных Triticum, извлеченной из UniProtKB (загружена в феврале 2018 г.). Параметрами поиска были: Окисление Met и пироглутаминовая кислота для N-terminus Gln в качестве вариабельных модификаций белка; допустимое значение массы 10 ppm (частей на миллион для ионов-предшественников и 0.01 Da для фрагментов MS/MS; протеолитический фермент не был выбран. Частота ложных срабатываний и вероятность обнаружения белка были рассчитаны с помощью валидатора соответствия спектра пептидов-мишеней (PSM), который работает в диапазоне от 0,01 до 0,05 для строгого и упрощенного поиска, соответственно. Данные трех повторных анализов LC-MS/MS были объединены. Количество пептидов определяли по количеству PSM и соответствующему количеству ионов.
Эпитопы Т-клеток были определены по их нативным последовательностям глиадина в том виде, в каком они были представлены до дезамидирования, поскольку лечение tTG2 не было включено в текущее исследование. Ожидается, что эти пептиды будут модифицированы in vivo собственной пластинкой tTG2 и станут иммуногенными, поскольку tTG2 дезамидирует глутамин (Q) в глутамат (E). Только дезамидированные пептиды помещаются в пептидсвязывающую канавку HLA-DQ2.5/8 и стимулируют CeD, стимулирующие CD4+ Т-клетки [23,24].
Результаты
Содержание белка варьировалось примерно от 8,2% до 11%, и во всех древних сортах пшеницы содержание белка превышало 10% (Таблица 2). Содержание крахмала было в пределах нормы (от 55% до 66%) для всех сортов пшеницы, а масса тысячи зерен (TKW) варьировалась от 30 до 41 г для разных сортов пшеницы.
При переваривании образцов каши с соками ЖКТ человека в условиях Ex vivo было получено множество фрагментов белка глютена, которые были идентифицированы с помощью HPLC-ESI MS/MS и программного обеспечения. В целом, для каждого образца было идентифицировано от 1051 до 2689 пептидов. Был составлен список уникальных пептидных последовательностей, не являющихся избыточными, который использовался для идентификации Т-клеточно-реактивных эпитопов в перечне [19]. В то время как при переваривании спельты высвобождается мало пептидов, их количество постепенно увеличивается у однозернянки (диплоидного) и эммер (тетраплоидного), а затем и у распространенных гексаплоидных сортов пшеницы Fram, Mirakel и Bastian. Таким образом, последние две разновидности имели наибольшее количество уникальных пептидов, из которых 144 и 155 были иммуногенными пептидами, содержащими Т-клеточные эпитопы (IPs), соответственно (Рисунок 1). Это контрастирует с показателями однозернянки, спельты и эммер, которые выпустили только 38, 45 и 68 IPs, соответственно (Рисунок 1). Таким образом, при переваривании однозернянки высвобождается значительно меньше IPs (18% пептидов), чем при переваривании Bastian (37% пептидов). Интересно, что ни один из переваренных сортов пшеницы не содержал неповрежденного – 33х, -26и или -25и мерного глиадина (Рисунок 2).
Хотя немногие пептиды идеально соответствовали Т-клеточно-реактивным эпитопам из перечня [19], олее крупные пептиды содержали эти IP-последовательности. Переваривание всех видов пшеницы содержало пептиды с эпитопами DQ2.5-glia-γ4c (QQPEQPFPQ), DQ2.5-glia-γ5 (QQPFPQQPQ) и DQ2.5-glia-ω1 (PFPQPQQPF), в то время как только гексаплоидные (T. aestivum) виды пшеницы, (Fram, Børsum, Bastian и Mirakel) высвобождали пептиды с эпитопами DQ2.5-glia-α2 (PQPELPYPQ) и DQ2.5-glia-α1b (PYPQPQLPY) (Рисунок 3). В частности, у однозернянки было обнаружено меньшее количество IPs с меньшей вариабельностью, поскольку были обнаружены только шесть из 22 Т-клеточно-реактивных эпитопов. Это в отличие от спельты, Bastian и Mirakel, которые выделяли 8, 11 и 13 различных Т-клеточно-реактивных эпитопов соответственно. Однако эпитопы DQ2.5-hor-2 (PQPQQPFPQ) и DQ2.5-glia-γ4b (PQPQQQFPQ) доминировали в родовых образцах пшеницы, поскольку они присутствовали только в одном и двух пептидах из обычных видов пшеницы соответственно.
γ-глиадины высвобождают наибольшее количество пептидов, содержащих Т-клеточные эпитопы, во всех типах пшеницы, как показано на Рисунке 4. Низкомолекулярные глютенины были вторым по величине источником IPs в однозернянке и эммер; ω-глиадины занимали второе место по величине, за ними следовали α-глиадины в распространенных гексаплоидных сортах пшеницы. Только Fram и Mirakel выпустили несколько IPS из высокомолекулярных глютенинов. Кроме того, мы также обнаружили содержащие Т-клеточные эпитопы пептиды из пшеничных секалинов во всех типах пшеницы, которые являются белками, обычно встречающимися во ржи и, скорее всего, идентифицируемыми здесь по гомологии.
Обсуждение
Целью данного исследования было охарактеризовать эпитопы Т-клеток, ассоциированные с CeD, в пептидах, выделяющихся при переваривании в желудочно-кишечном тракте ex vivo трех древних сортов пшеницы (однозернянка, эммер и спельта), и 4х обычных норвежских сортов пшеницы (Fram, Børsum, Bastian и Mirakel). Аналогично работе Shan и соавт. [10], в нескольких исследованиях было установлено, что устойчивые к перевариванию 33мерные (LQLQPFPQPQLPYPQPQLPYPQPQLPYPQPQPF), и 25мерные (LGQQQPFPPQQPYPQPQPFPSQQPY) фрагменты из α-глиадина [25], и 26мерные (FLQPQQPFPQQPQQPYPQQPQQPFPQ) из γ-глиадина [26] содержат большинство Т-клеточно-реактивных эпитопов, участвующих в иммунной реакции CeD [27]. Удивительно, но эти крупные иммунодоминантные пептиды были расщеплены на более мелкие, но все же содержащие Т-клеточные эпитопы пептиды современными пищеварительными системами ex vivo. Это контрастирует с предыдущими экспериментами по расщеплению in vitro, показавшими, что 33-мерные, 26-мерные и 25-мерные пептиды были устойчивы к расщеплению. В то время как в предыдущих моделях пищеварения использовались пищеварительные ферменты крупного рогатого скота или свиньи, в нынешних экспериментах использовались гастродуоденальные аспираты человека, что повышает физиологическую значимость модели и, следовательно, более точно имитирует желудочно-кишечное пищеварение in vivo [28]. Однако, поскольку мы использовали гастродуоденальные аспираты от здоровых людей, у которых, по-видимому, не было целиакии, мы не можем исключить возможность того, что пациенты с целиакией переваривают глютен по-разному либо из-за генетических различий в кишечных пищеварительных ферментах, либо из-за различий в переваривании глютена с помощью микробов.
Переработанные пшеничные пептиды содержали как одиночные, так и множественные перекрывающиеся иммуногенные последовательности ядер. В то время как однозернянка выделяла наименьшее количество возможных Ips, а за ней следовала спельта, Bastian выделил наибольшее количество IPs. Хотя процентное соотношение IPs к общему количеству пептидов существенно не различалось между сортами пшеницы с разными геномами, у древних сортов, как правило, процентное соотношение IPs было ниже. Это противоречит выводам Prandi и соавт. [29], которые сообщили, что при культивировании in vitro старых сортов пшеницы образуется больше IPs по сравнению с современными сортами. Однако их современные образцы пшеницы включали как однозернянку, так и спельту, которые в нашем исследовании были классифицированы как древние, а их старые сорта включали только тетраплоидные и гексаплоидные сорта пшеницы (T. aestivum L., T. turgidum var. durum Desf., T. turgidum var. dicoccum L (эммер)), что затрудняет сравнение результатов. Наши результаты также противоречат выводам Malalgoda и соавт. [30], которые не обнаружили различий между древними и современными сортами пшеницы в пептидах, содержащих Т-клеточные эпитопы, высвобождающихся после расщепления глиадина в геле свиным химотрипсином. Однако эта система пищеварения в геле с коммерческими ферментами не имитирует пищеварение в ЖКТ человека так, как это делает современная пищеварительная система ex vivo, использующая гастродуоденальные соки человека.
Локус гена 33х-мерного α-глиадина расположен на хромосоме 6D только у сортов гексаплоидной пшеницы [31]. Таким образом, последовательность 33mer отсутствует в однозернянке (AA) и эммер (AABB) но может присутствовать в спельте, Fram, Børsum, Bastian и Mirakel (AABBDD). Пеприд 33х-мерного α-глиадина содержит шесть перекрывающихся последовательностей Т-клеточных эпитопов: одну копию DQ2.5--связывающего глиадин пептида, glia-α1a (PFPQPQLPY), две копии DQ2.5-glia-α1b (PYPQPQLPY) и три копии DQ2.5-glia-α2 (PQPQLPYPQ). В то время как ни один из этих эпитопов не был обнаружен в диплоидной однозернянке, переваренные эммер и спельта выделили один пептид с эпитопом DQ2.5-glia-α1a. Напротив, эти Т-клеточные эпитопы присутствовали во многих пептидах гексаплоидных сортов пшеницы. В частности, Mirakel, Bastian и Børsum выделили несколько 33-мерных фрагментов (Рисунок 2). Глиадины, полученные из хромосомы 6D, расщеплялись в разных положениях, образуя пептиды разной длины со сходными или множественными перекрывающимися последовательностями эпитопов. Таким образом, при переваривании этих сортов пшеницы высвобождается больше пептидов, содержащих Т-клеточные эпитопы, чем у диплоидных и тетраплоидных сортов пшеницы, у которых отсутствует хромосома 6D.
26-мерный пептид, полученный из γ-глиадина, содержит два перекрывающихся Т-клеточных эпитопа: DQ2.5-glia-γ3, (QQPQQPYPQ) и DQ2.5-glia-γ4c (QQPQQPFPQ). В то время как эпитоп DQ2.5-glia-γ3 отсутствовал в однозернянке и спельте, н присутствовал в продуктах из других гексаплоидных сортов пшеницы и эммер. Хотя эпитоп DQ2.5-glia-γ4c был наиболее доминирующим пептидным эпитопом во всех переваренных образцах пшеницы, его содержание варьировалось от умеренного в однозернянке и спельте до почти трехкратного увеличения в обычных сортах пшеницы. Другие доминирующие эпитопы, DQ2.5-glia-γ5 (QQPFPQQPQ) и DQ2.5-glia-ω1 (PFPQPQQPF), а также DQ2.5-glia-γ4c, были обнаружены после комплексного протеолитического расщепления глютена с использованием методов выделения человеческих моноклональных антител для идентификации эпитопов [32].
25-мерный пептид α-глиадина не был обнаружен во всех образцах. Однако однозернянка, эммер, Børsum, Bastian и Mirakel, но не спельта и Fram, выпустили укороченный 20ти-мерный пептид (31LGQQQPFPPQQPYPQPQPFPS51). Несколько исследований показали, что 13ти-мерная последовательность 31LGQQQPFPPQQPY43 в составе 25-мерного пептида α-глиадина и укороченного 20-мерного пептида, обнаруженная в этом исследовании, активирует врожденную иммунную систему [33] путем усиления регуляции интерлейкина-15, циклооксигеназы-2 (COX-2), экспрессии CD25 и CD83 на собственной пластинке макрофагов, моноцитах, и дендритных клеток перед любой стимуляцией CD4+ Т-клеток [34]. Совсем недавно Barone и соавт. [35] показали, что 13ти-мерные индуцировали изменение везикулярного обмена в эпителиальной клеточной линии рака толстой кишки Caco-2. Это приводит к сверхэкспрессии транс-представленного комплекса IL-15/IL5R который индуцирует EGFR-зависимую пролиферацию клеток, что может объяснить ремоделирование слизистой оболочки при CeD. Таким образом, врожденная иммунная система также может быть вовлечена в этот аспект патогенеза CeD.
Выводы
При переваривании диплоидной, тетраплоидной и гексаплоидной пшеницы в желудочно-кишечном тракте человека ex vivoобразуются пептиды, содержащие Т-клеточные эпитопы, в зависимости от геномных особенностей. Переваривание пшеницы желудочно-кишечными соками человека привело к иной схеме расщепления белка по сравнению с ранее проведенными исследованиями, в которых использовались ферменты свиного или бычьего происхождения. В нашем исследовании иммунодоминантные пептиды (33-мерные и 25-мерные α-глиадиновые пептиды и 26-мерный γ-глиадиновый пептид) были обнаружены не в их интактной форме, а в виде разрушенных фрагментов. Существующая модель пищеварения не включала разложение и всасывание в слизистой оболочке. Следовательно, биодоступность и способность этих пептидов достигать собственной пластинки и связываться с tTG2 посредством дезамидирования и, таким образом, приобретать свойства связывания с HLA-DQ2.5/8 и превращаться в Т-клеточные эпитопы, еще предстоит изучить. Тем не менее, результаты показали, что древние виды пшеницы могут быть менее токсичными в плане CeD по сравнению с обычными гексаплоидными сортами. Вопрос о том, может ли более широкое использование этих наследственных вариантов пшеницы у генетически предрасположенных людей снизить CeD нуждается в дальнейшей оценке, но частота диагностируемых CeD в детском возрасте была связана с количеством глютена в рационе [36].
Вклад Авторов: G.E.V., T.G.D., A.K.U. и T.A. разработали исследование; A.R. провел аспирацию желудка и двенадцатиперстной кишки у добровольцев; R.R., G.M. и G.P. провели анализы; R.R. и T.A. проанализировали данные и написали рукопись; T.A., R.R., G.M., G.P., T.G.D., G.E.V., A.R., T.E.L., T.S.H., P.F. и A.K.U. внесли свой вклад в интерпретацию результатов и пересмотр рукописи. Все авторы ознакомились с опубликованной версией рукописи и согласились с ней.
Финансирование: Данное исследование не получало внешнего финансирования.
Конфликт интересов: Авторы заявляют об отсутствии конфликта интересов.
Источники
1. Lohi, S.; Mustalahti, K.; Kaukinen, K.; Laurila, K.; Collin, P.; Rissanen, H.; Lohi, O.; Bravi, E.; Gasparin, M.; Reunanen, A.; et al. Increasing prevalence of coeliac disease over time. Aliment. Pharmacol. Ther. 2007, 26, 1217–1225. [CrossRef] [PubMed]
2. Green, P.H.; Cellier, C. Celiac disease. New Engl. J. Med. 2007, 357, 1731–1743. [CrossRef] [PubMed]
3. Katz, K.D.; Rashtak, S.; Lahr, B.D.; Melton, J.L.I.; Krause, P.K.; Maggi, K.; Talley, N.J.; Murray, J.A. Screening for celiac disease in a north american population: Sequential serology and gastrointestinal symptoms. Am. J. Gastroenterol. 2011, 106, 1333–1339. [CrossRef] [PubMed]
4. Schalk, K.; Lexhaller, B.; Koehler, P.; Scherf, K.A. Isolation and characterization of gluten protein types from wheat, rye, barley and oats for use as reference materials. PLoS ONE 2017, 12, e0172819. [CrossRef] [PubMed]
5. Kagnoff, M.F. Celiac disease: Pathogenesis of a model immunogenetic disease. J. Clin. Investig.
2007, 117, 41–49. [CrossRef]
6. Sollid, L.M. Coeliac disease: Dissecting a complex inflammatory disorder. Nat. Rev. Immunol. 2002, 2, 647. [CrossRef]
7. Karell, K.; Louka, A.S.; Moodie, S.J.; Ascher, H.; Clot, F.; Greco, L.; Ciclitira, P.J.; Sollid, L.M.; Partanen, J.; Members of the European Genetics Cluster on Celiac Disease. HLA types in celiac disease patients not carrying the DQA1* 05-DQB1* 02 (DQ2) heterodimer: Results from the European Genetics Cluster on Celiac Disease. Hum. Immunol. 2003, 64, 469–477. [CrossRef]
8. Trynka, G.; Hunt, K.A.; Bockett, N.A.; Romanos, J.; Mistry, V.; Szperl, A.; Bakker, S.F.; Bardella, M.T.; Bhaw-Rosun, L.; Castillejo, G.; et al. Dense genotyping identifies and localizes multiple common and rare variant association signals in celiac disease. Nat. Genet. 2011, 43, 1193. [CrossRef]
9. Shewry, P.R.; Halford, N.G.; Lafiandra, D. Genetics of wheat gluten proteins. Adv. Genet. 2003, 49, 111–184.
10. Shan, L.; Molberg, Ø.; Parrot, I.; Hausch, F.; Filiz, F.; Gray, G.M.; Sollid, L.M.; Khosla, C. Structural basis for gluten intolerance in celiac sprue. Science 2002, 297, 2275–2279. [CrossRef]
11. Stepniak, D.; Vader, L.W.; Kooy, Y.; van Veelen, P.A.; Moustakas, A.; Papandreou, N.A.; Eliopoulos, E.; Drijfhout, J.W.; Papadopoulos, G.K.; Koning, F. T-cell recognition of HLA-DQ2-bound gluten peptides can be influenced by an N-terminal proline at p-1. Immunogenetics 2005, 57, 8–15. [CrossRef] [PubMed]
12. Arentz-Hansen, H.; Körner, R.; Molberg, Ø.; Quarsten, H.; Vader, W.; Kooy, Y.M.; Lundin, K.E.A.; Koning, F.; Roepstorff, P.; Sollid, L.M.; et al. The intestinal T cell response to α-gliadin in adult celiac disease is focused on a single deamidated glutamine targeted by tissue transglutaminase. J. Exp. Med. 2000, 191, 603–612. [CrossRef] [PubMed]
13. Sjostrom, H.; Lundin, K.E.; Molberg, O.; Korner, R.; McAdam, S.N.; Anthonsen, D.; Quarsten, H.; Norén, O.; Roepstorff, P.; Thorsby, E.; et al. Identification of a gliadin T-cell epitope in coeliac disease: General importance of gliadin deamidation for intestinal T-cell recognition. Scand. J. Immunol. 1998, 48, 111–115. [CrossRef]
14. Gianfrani, C.; Camarca, A.; Mazzarella, G.; Di Stasio, L.; Giardullo, N.; Ferranti, P.; Picariello, G.; Aufiero, V.R.; Picascia, S.; Troncone, R.; et al. Extensive in vitro gastrointestinal digestion markedly reduces the immune-toxicity of triticum monococcum wheat: Implication for celiac disease. Mol. Nutr. Food Res. 2015, 59, 1844–1854. [CrossRef]
15. Huang, S.; Sirikhachornkit, A.; Su, X.; Faris, J.; Gill, B.; Haselkorn, R.; Gornicki, P. Genes encoding plastid acetyl-CoA carboxylase and 3-phosphoglycerate kinase of the Triticum/Aegilops complex and the evolutionary history of polyploid wheat. Proc. Natl. Acad. Sci. USA 2002, 99, 8133–8138. [CrossRef] [PubMed]
16. Li, A.; Liu, D.; Yang, W.; Kishii, M.; Mao, L. Synthetic hexaploid wheat: Yesterday, today, and tomorrow.
Engineering 2018, 4, 552–558. [CrossRef]
17. Wieser, H. Comparative investigations of gluten proteins from different wheat species. III. N-terminal amino acid sequences of α-gliadins potentially toxic for coeliac patients. Eur. Food Res. Technol. 2001, 213, 183–186.
18. Deng, Y.; Gruppen, H.; Wierenga, P.A. Comparison of protein hydrolysis catalyzed by bovine, porcine, and
human trypsins. J. Agric. Food Chem. 2018, 66, 4219–4232. [CrossRef]
19. Sollid, L.M.; Qiao, S.-W.; Anderson, R.P.; Gianfrani, C.; Koning, F. Nomenclature and listing of celiac disease relevant gluten T-cell epitopes restricted by HLA-DQ molecules. Immunogenetics 2012, 64, 455–460. [CrossRef]
20. McCleary, B.; Gibson, T.; Solah, V.; Mugford, D. Total starch measurement in cereal products: Interlaboratory evaluation of a rapid enzymic test procedure. Cereal Chem. 1994, 71, 501–504.
21. Ulleberg, E.K.; Comi, I.; Holm, H.; Herud, E.B.; Jacobsen, M.; Vegarud, G.E. Human gastrointestinal juices intended for use in in vitro digestion models. Food Dig. 2011, 2, 52–61. [CrossRef] [PubMed]
22. Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [CrossRef] [PubMed]
23. Molberg, Ø.; Mcadam, S.N.; Körner, R.; Quarsten, H.; Kristiansen, C.; Madsen, L.; Fugger, L.; Scott, H.; Norén, O.; Roepstorff, P.; et al. Tissue transglutaminase selectively modifies gliadin peptides that are recognized by gut-derived T cells in celiac disease. Nat. Med. 1998, 4, 713. [CrossRef] [PubMed]
24. van de Wal, Y.; Kooy, Y.; van Veelen, P.; Peña, S.; Mearin, L.; Papadopoulos, G.; Koning, F. Cutting edge: Selective deamidation by tissue transglutaminase strongly enhances gliadin-specific T cell reactivity.
J. Immunol. 1998, 161, 1585–1588.
25. Ciccocioppo, R.; Di Sabatino, A.; Corazza, G.R. The immune recognition of gluten in coeliac disease.
Clin. Exp. Immunol. 2005, 140, 408–416. [CrossRef]
26. Mamone, G.; Ferranti, P.; Rossi, M.; Roepstorff, P.; Fierro, O.; Malorni, A.; Addeo, F. Identification of a peptide from α-gliadin resistant to digestive enzymes: Implications for celiac disease. J. Chromatogr. B 2007, 855, 236–241. [CrossRef]
27. Gianfrani, C.; Siciliano, R.A.; Facchiano, A.M.; Camarca, A.; Mazzeo, M.F.; Costantini, S.; Salvati, V.M.; Maurano, F.; Mazzarella, G.; Iaquinto, G.; et al. Transamidation of wheat flour inhibits the response to gliadin of intestinal T cells in celiac disease. Gastroenterology 2007, 133, 780–789. [CrossRef]
28. Asledottir, T.; Picariello, G.; Mamone, G.; Ferranti, P.; Roseth, A.; Devold, T.G.; Vegarud, G.E. Degradation of beta-casomorphin-7 through in vitro gastrointestinal and jejunal brush border membrane digestion.
J. Dairy Sci. 2019, 102, 8622–8629. [CrossRef]
29. Prandi, B.; Tedeschi, T.; Folloni, S.; Galaverna, G.; Sforza, S. Peptides from gluten digestion: A comparison between old and modern wheat varieties. Food Res. Int. 2017, 91, 92–102. [CrossRef]
30. Malalgoda, M.; Meinhardt, S.W.; Simsek, S. Detection and quantitation of immunogenic epitopes related to celiac disease in historical and modern hard red spring wheat cultivars. Food Chem. 2018, 264, 101–107. [CrossRef]
31. Molberg, Ø.; Uhlen, A.K.; Sollid, L.M.; Jensen, T.; Flæte, N.S.; Arntz-Hansen, H.; Raki, M.; Lundin, K.E.A.; Sollid, L.M. Mapping of gluten T-cell epitopes in the bread wheat ancestors: Implications on celiac disease. Gastoenterology 2005, 128, 393–401. [CrossRef] [PubMed]
32. Dorum, S.; Steinsbo, O.; Bergseng, E.; Arntzen, M.O.; de Souza, G.A.; Sollid, L.M. Gluten-specific antibodies of celiac disease gut plasma cells recognize long proteolytic fragments that typically harbor T-cell epitopes. Sci Rep. 2016, 6, 25565. [CrossRef] [PubMed]
33. Hüe, S.; Mention, J.-J.; Monteiro, R.C.; Zhang, S.; Cellier, C.; Schmitz, J.; Verkarre, V.; Fodil, N.; Bahram, S.; Cerf-Bensussan, N.; et al. A direct role for NKG2D/MICA interaction in villous atrophy during celiac disease. Immunity 2004, 21, 367–377. [CrossRef] [PubMed]
34. Maiuri, L.; Ciacci, C.; Ricciardelli, I.; Vacca, L.; Raia, V.; Auricchio, S.; Picard, J.; Osman, M.; Quaratino, S.; Londei, P.M. Association between innate response to gliadin and activation of pathogenic T cells in coeliac disease. Lancet 2003, 362, 30–37. [CrossRef]
35. Barone, M.V.; Zanzi, D.; Maglio, M.; Nanayakkara, M.; Santagata, S.; Lania, G.; Miele, E.; Ribecco, M.T.S.; Maurano, F.; Auricchio, R.; et al. Gliadin-mediated proliferation and innate immune activation in celiac disease are due to alterations in vesicular trafficking. PLoS ONE 2011, 6, e17039. [CrossRef]