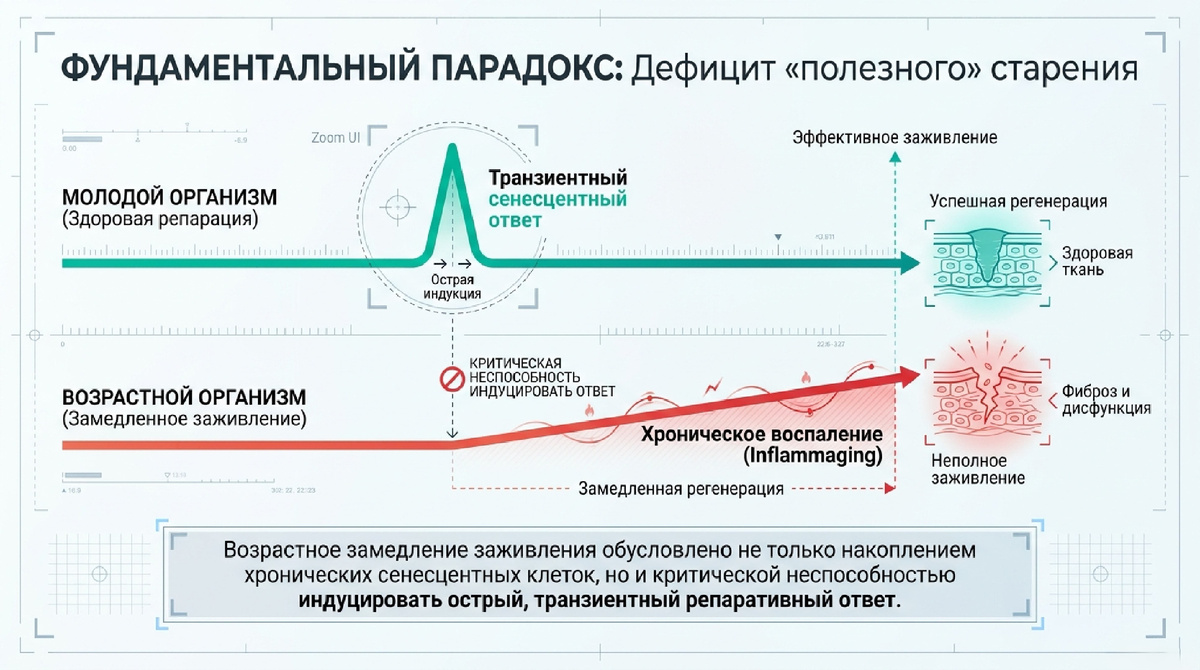
1. Введение: Роль транзиторного старения в регенерации тканей
В современной молекулярной геронтологии происходит фундаментальный пересмотр парадигмы клеточного старения (senescence). Для разработки эффективных биомедицинских стратегий критически важно проводить разграничение между накоплением «хронических» сенильных клеток, характерным для стареющих тканей, и «острым» (транзиторным) старением, которое является эволюционно консервативным механизмом успешного восстановления.
Временная активация программы старения в ответ на повреждение в молодом возрасте выступает не как признак дегенерации, а как необходимый физиологический триггер. Через специфический секреторный фенотип (SASP), включающий факторы роста и матриксные металлопротеиназы, эти клетки координируют архитектурную перестройку ткани. Нарушение этой динамики в процессе старения организма — «функциональный захват» транзиторной программы старения хроническими воспалительными процессами — является центральным фактором снижения регенеративного потенциала.
2. Сравнительный анализ кинетики закрытия ран
Анализ макроскопических данных молодых (2 месяца) и старых (24 месяца) мышиных моделей демонстрирует существенную деградацию темпов репарации у пожилых особей. Расхождение траекторий заживления становится статистически и клинически значимым на ранних этапах.
Ключевые дифференциаторы процесса заживления:
- День 6 (Ранний этап): Начало выраженной задержки. У старых моделей площадь открытой раны значительно превышает показатели молодой группы.
- День 12–15: Усугубление дефицита эпителизации. В то время как молодые особи демонстрируют активную контракцию раневого ложа, у пожилых сохраняется обширный тканевый дефект.
- День 18 (Критическая точка): Почти все молодые мыши (100%) достигают полного закрытия ран. В группе старых моделей этот показатель составляет критически низкие 5,9%.
- День 24 (Конечная точка): Несмотря на продолжительный период наблюдения, полного заживления достигают лишь 88,2% старых особей.
Данная задержка указывает на глубокую дезорганизацию молекулярного ответа, при которой старение организма не просто замедляет, а качественно искажает регенеративный каскад.
3. Динамика маркеров клеточного старения в раневой ткани
Успех регенерации определяется способностью ткани сформировать координированный пик клеточного старения. В молодом возрасте наблюдается четкая временная индукция маркеров, в то время как у старых особей этот ответ аттенуирован на фоне повышенного «фонового» уровня хронического старения.
Сравнительная экспрессия биомаркеров старения
Маркер / Группа / Норма (Day 0) / Пиковая точка (День) / Динамика в ране
Cdkn2a (p16) / Молодые / Низкая / День 6 (высокая) / Мощная транзиторная индукция
Cdkn2a (p16) / Старые / Повышенный фон / - / Отсутствие подъема выше базы
Cdkn1a (p21) / Молодые / Низкая / День 18 / Четкий пик (иммунофлуоресценция)
Cdkn1a (p21) / Старые / Умеренная / - / Отсутствие значимого отклика
SA-β-gal / Молодые / <10% / День 6–12 (>20%) / Резкое временное накопление
Cdkn1a (p21) / Старые / ~10% / - / Инертность к ранению
Важным качественным дифференциатором является интенсивность сигнала: анализ на уровне единичных клеток показал, что молодые сенильные фибробласты демонстрируют на 34% более высокую экспрессию Cdkn2a на клетку по сравнению с клетками старых особей. Таким образом, старые раны характеризуются не только меньшим количеством сенильных клеток, но и более «слабым», функционально дефектным профилем старения.
4. Транскриптомный атлас раневого ложа (scRNA-seq)
Секвенирование единичных клеток на 6-й день выявило 20 гетерогенных кластеров. Анализ подтверждает, что фибробласты являются доминирующей популяцией, вступающей в состояние старения в ране (популяция p16+/p21+/Ki67–).
Анализ клеточных популяций:
- Дефицит сенильной популяции: В молодых ранах доля клеток p16+/p21+/Ki67– составляет 4,1%, сокращаясь у старых мышей до 1,7%.
- Критическая потеря Cluster 15: Идентифицировано значительное сокращение специфического Кластера 15 (ECM-продуцирующие фибробласты) у пожилых особей. Эта популяция является ключевым звеном в производстве матрикса.
- Архитектурные последствия: Снижение численности этих клеток ведет к разрушению регуляторного центра раны. В молодом организме эти клетки выступают «дирижерами» процесса, тогда как их дефицит в старой ткани приводит к деградации сигнальных связей, необходимых для морфогенеза.
5. Функциональная сигнатура фибробластов: ECM-профиль против воспаления
Центральным открытием является качественный сдвиг транскрипционного профиля сенильных фибробластов: от «ремоделирования матрикса» у молодых к «стабилизации и воспалению» у старых.
Молодые фибробласты (Про-репаративный фенотип)
Ориентированы на динамическое перестроение (rebuilding) матрикса:
- Гены матрикса: Высокая экспрессия фибриллярных коллагенов (Col1a1, Col1a2, Col6a1-3) и металлопротеиназ (Mmp2, Mmp3, Mmp11, Mmp14), обеспечивающих пластичность.
- Матрицеллярный сигналинг: Секреция факторов роста и миграции (Igfbp2, Hgf, Ereg).
- Коммуникация: Согласно CellChat, эти клетки формируют мощный фибробласто-центричный коммуникационный хаб, транслируя сигналы через пути коллагена, тенасцина и тромбоспондина другим фибробластам и иммунным клеткам.
Старые фибробласты (Про-воспалительный фенотип)
Демонстрируют признаки протеостатического стресса и хронического воспаления:
- Сдвиг SASP: Активация интерферонового сигналинга (interferon signaling) и рибосомального стресса. Усиление секреции Il6 и хемокинов (Cxcl1, 2, 3, 5, 10), поддерживающих состояние inflammaging.
- Матриксная стабилизация: Вместо фибриллярных белков клетки переключаются на коллагены базальной мембраны и адгезивные молекулы (Col4a2, Col5a1, Col5a2, Col12a1, Col14a1). Это ведет к патологической стабилизации матрикса и избыточной адгезии (через ламинин и фибронектин), что блокирует нормальное разрешение раны.
- Коллапс коммуникации: Происходит разрушение сигнальных ребер Senescent_FB → Other_FB. Иммунные клетки становятся основными отправителями сигналов, что характерно для хронического воспалительного процесса, а не регенерации.
6. Трансляционная значимость и человеческая валидация
Данные мышиных моделей были полностью подтверждены анализом scRNA-seq ран человека (Liu et al., 2025). У молодых доноров (20+ лет) на 7-й день после ранения зафиксирована аналогичная популяция транзиторных сенильных фибробластов со статусом p16+/p21+/MKI67–.
Эти клетки локализуются преимущественно в Кластере 2 (Активированные фибробласты) и демонстрируют идентичную молодым мышам сигнатуру: активацию программ созревания коллагена и его гликозилирования. Это подтверждает, что острый сенильный ответ является универсальным и консервативным механизмом эффективного заживления, утрата которого ведет к патологии.
7. Заключение и обоснование биомедицинских разработок
Задержка заживления при старении обусловлена двойным дефектом: количественным дефицитом про-репаративных сенильных клеток и качественным вырождением их секреторного профиля в сторону протеотоксического стресса и воспаления.
Стратегические направления разработок:
- Модуляция секреторного фенотипа (SASP): Перепрограммирование секрета старых клеток для подавления интерферонового ответа и восстановления экспрессии факторов роста (Igfbp2, Hgf).
- Восстановление транзиторной индукции: Применение молекулярных триггеров для стимуляции нормального входа фибробластов в состояние «полезного» старения в раннем окне (6–7 день) у пожилых пациентов.
- Таргетное управление популяциями: Использование сенолитиков, таких как ABT-263, требует строгого тайминга. Данные подтверждают, что удаление базовых «хронических» клеток препаратом ABT-263 до нанесения раны улучшает последующее заживление, очищая микросреду.
Финальная рекомендация: Терапия должна быть направлена на восстановление баланса между удалением хронического сенильного фона и сохранением/стимуляцией острых сенильных популяций. Неселективное применение сенолитиков в момент ранения может быть контрпродуктивным, подавляя естественные механизмы регенерации.