1. Введение: Кишечный барьер в контексте системного старения
Кишечный эпителий представляет собой крупнейший в организме эндокринный и иммунный интерфейс, определяющий не только локальный гомеостаз, но и системное долголетие. Его барьерная функция, обеспечиваемая комплексами плотных и адгезионных контактов, критически важна для предотвращения транслокации люминальных антигенов и патогенов. С возрастом этот интерфейс претерпевает деградацию, характеризующуюся повышением проницаемости («leaky gut») и развитием инфламейджинга — хронического системного воспаления низкой интенсивности, являющегося драйвером метаболических и нейродегенеративных расстройств.
Традиционно старение кишечника рассматривалось как описательный феномен накопления структурных дефектов. Однако новейшее исследование Zhang et al. (2026) радикально меняет эту парадигму, идентифицируя ион магния (Mg²⁺) как «мастер-регулятор» целостности эпителия. Понимание магниевого гомеостаза позволяет перейти от фиксации возрастных изменений к их механистическому анализу: дефицит Mg²⁺ выступает не просто как нутритивный маркер, а как первичный молекулярный триггер перестройки протеома и дестабилизации клеточных контактов. Клиническая значимость этого открытия заключается в возможности таргетной нутритивной коррекции процессов, которые ранее считались неизбежным следствием биологического износа.
2. Возрастная динамика содержания магния и механизмы его транспорта
Анализ методом масс-спектрометрии с индуктивно-связанной плазмой (ICP-MS) демонстрирует, что тканеспецифическое снижение уровня Mg²⁺ является универсальным признаком старения у различных видов:
- Приматы (Cynomolgus monkeys): Уровни Mg²⁺ в толстой кишке демонстрируют выраженное возраст-зависимое снижение, причем у самок этот процесс протекает более интенсивно.
- Человек: Концентрация Mg²⁺ в дистальных отделах кишечника отрицательно коррелирует с биологическим возрастом (выборка 22–91 год) у представителей обоих полов.
- Мыши: Возрастное снижение магния локализовано преимущественно в толстой кишке, тогда как в тонком кишечнике и других органах уровень остается относительно стабильным.
Тканеспецифичность содержания Mg²⁺ при старении (Z-score на основе ICP-MS):
Орган / Состояние Mg²⁺ при старении / Статистическая значимость
Толстая кишка / Значительное снижение / p < 0.001
Головной мозг / Стабильно / NS
Сердце / Стабильно / NS
Печень / Стабильно / NS
Почки / Стабильно / NS
Костная ткань / Нет значимых изменений / NS
Критическим фактором этого снижения является нарушение работы транспортных систем. С возрастом наблюдается подавление экспрессии эпителиального канала TRPM6 при относительной стабильности его гомолога TRPM7, что указывает на селективную деградацию механизмов абсорбции. Параллельное снижение экспрессии SLC41A3 коррелирует с падением внутритканевого уровня Mg²⁺.
Особого внимания заслуживает роль экспортера CNNM4. В отличие от каналов всасывания, снижение уровня CNNM4 в стареющем кишечнике и при язвенном колите может представлять собой компенсаторную реакцию: эпителиоциты ограничивают базалатеральный экспорт Mg²⁺ в системный кровоток, пытаясь сохранить критический уровень иона внутри клетки для поддержания минимальной ферментативной активности. Однако этой адаптации оказывается недостаточно для предотвращения протеомного ремоделирования.
3. Ремоделирование фосфопротеома: дестабилизация адгезионных контактов
Поскольку Mg²⁺ является незаменимым кофактором для связывания Mg·ATP в каталитических центрах киназ, его локальный дефицит искажает ландшафт фосфорилирования. Гипомагниемия инициирует масштабную перестройку 1672 фосфосайтов, что напрямую ведет к дестабилизации адгезионных комплексов, включая белки CTNNA1 (α-катенин), NF2 (мерлин) и TJP2.
Ключевым молекулярным событием является снижение фосфорилирования сайта S986 (у человека) / S968 (у мышей) белка плотных контактов TJP2. Исследования с использованием фосфомиметической модификации TJP2 S986D (замена серина на аспаргиновую кислоту) подтвердили, что восстановление фосфостатуса этого сайта способно реанимировать экспрессию барьерных белков (JAM1, Claudin-1) и замедлить клеточный сенесценс.
Дисрегуляция киназной активности носит двунаправленный характер. С одной стороны, дефицит Mg·ATP подавляет анаболические пути mTOR/ERK2, критически важные для регенерации. С другой стороны, сопутствующее нарушение гомеостаза кальция и стрессовые сигналы активируют киназы CDK3 и SRPK1, способствующие клеточному старению. Таким образом, гипомагниемия «перепрограммирует» фосфопротеом, лишая эпителий структурной гибкости и способности к самовосстановлению.
4. Перестройка N-гликопротеома и потеря структурной целостности
Синергетический эффект фосфопротеомного ремоделирования усиливается аберрантным N-гликозилированием. Магний модулирует активность гликозилтрансфераз и гликозидаз, и его дефицит приводит к синтезу белков с нарушенной гликановой «шубой», что фатально сказывается на их стабильности и взаимодействии.
Мультиомиксный анализ выявил специфические изменения N-гликанов в белках адгезии:
- Снижение маннозных типов: Например, в интегрин-альфа-6 (ITGA6, сайт N927), что ослабляет связь эпителия с базальной мембраной.
- Рост фукозилирования: В частности, повышение уровня фукозилированных гликанов в кадгерине-17 (CDH17, сайт N573), что характерно для воспалительных фенотипов.
Эти молекулярные дефекты при дефиците магния практически идентичны профилю естественного старения. Гликопротеомный анализ подтверждает: гипомагниемия является не просто маркером, а активным драйвером «гликанового старения», компрометирующим межклеточные взаимодействия и архитектуру крипт кишечника. Это подчеркивает особую уязвимость женского организма, где изменения PTM (фосфо- и глико-) в ответ на колебания уровня магния выражены более рельефно.
5. Фенотипические последствия: парадокс возраста и воспаление
Экспериментальные данные выявляют значительный контраст в ответе на дефицит Mg²⁺ в зависимости от возраста. У молодых мышей ограничение магния не приводит к немедленному сокращению колона или разрушению барьера. Это объясняется высокой регуляторной емкостью молодого организма: эффективная почечная реабсорбция и адаптивное повышение фракционного всасывания в кишечнике позволяют временно буферизировать дефицит. С возрастом эти компенсаторные механизмы угасают, делая кишечник беззащитным перед микронутриентным дисбалансом.
У пожилых животных гипомагниемия манифестирует ускоренным старением:
- Морфология: Выраженное сокращение длины толстой кишки и атрофия эпителия.
- Сенесценс: Резкий рост маркера p21.
- Иммунный профиль: Про-воспалительное ремоделирование с инфильтрацией макрофагов и ростом цитокинов (IL-6, IL-1β, KC) при снижении пула защитных эозинофилов.
В модели DSS-индуцированного колита дефицит магния критически снижает порог выживаемости, тогда как восполнение уровня Mg²⁺ восстанавливает целостность барьера и подавляет экспрессию p21, возвращая ткани черты «молодого» фенотипа.
6. Клиническая значимость: эпидемиологический анализ UK Biobank
Масштабное исследование когорты UK Biobank (n = 182 213) с 13-летним периодом наблюдения подтверждает фундаментальную роль магния в превентивной гастроэнтерологии. Анализ высшего и низшего квинтилей потребления выявил значительное защитное действие магния (Hazard Ratios):
- Болезнь Крона: HR = 0.495 (снижение риска более чем в два раза).
- Язвенный колит: HR = 0.632.
- Синдром раздраженного кишечника (СРК): HR = 0.786.
- Дивертикулярная болезнь: HR = 0.799.
Важно отметить, что для целиакии корреляция оказалась статистически незначимой (HR = 0.918, p=0.496), что подтверждает специфичность действия магния на механизмы барьерной адгезии и воспаления, а не на аутоиммунные процессы, опосредованные глютеном. Результаты оставались стабильными даже после исключения участников с признаками нутритивной недостаточности (по критериям GLIM), что доказывает: защитный эффект магния является самостоятельным фактором, не зависящим от общей калорийности рациона.
Терапевтическое окно: Оптимальный диапазон потребления для снижения риска кишечных патологий составляет 334.7 – 420.0 мг/день.
7. Заключение: Перспективы магниевой терапии в геронтологии
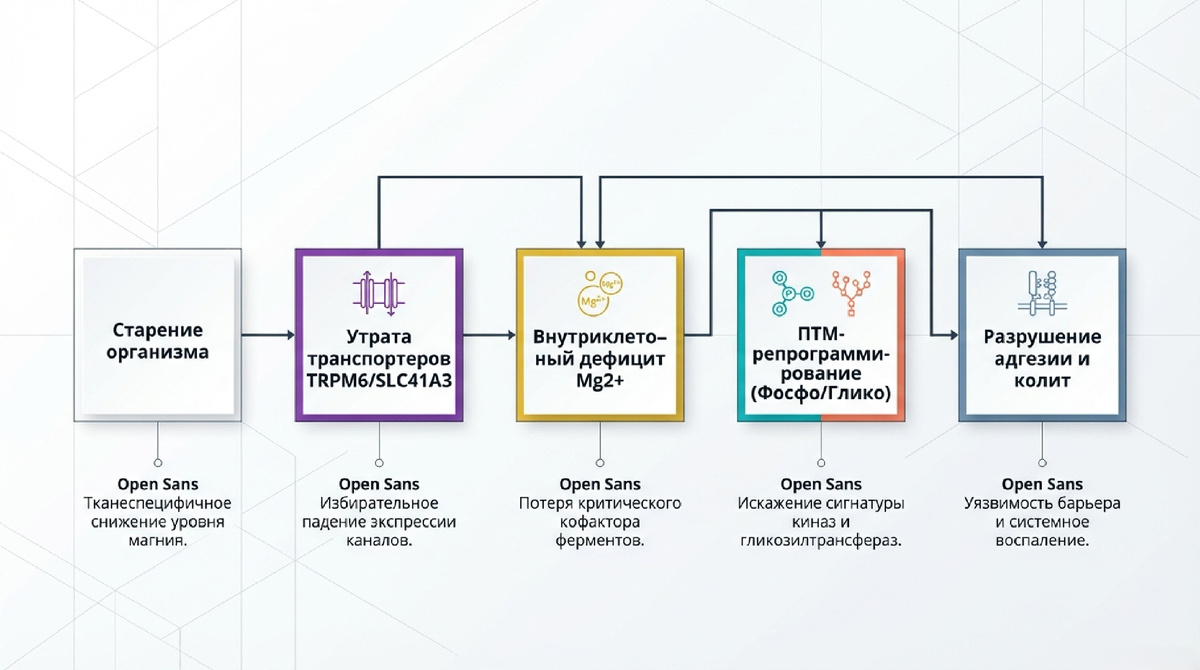
Интеграция мультиомиксных данных позволяет сформулировать завершенную концептуальную цепь патогенеза: [Возрастное снижение TRPM6/SLC41A3] → [Тканевый дефицит Mg²⁺] → [Дисрегуляция фосфо- и N-гликопротеома (TJP2, ITGA6, CDH17)] → [Разрушение адгезионных контактов] → [Ускоренное старение и инфламейджинг].
Статус магния должен рассматриваться не просто как элемент диетологии, а как критический биомаркер кишечной резистентности. Для клиницистов это означает необходимость регулярного мониторинга и коррекции магниевого статуса у пациентов среднего и пожилого возраста (особенно у женщин) как базовой стратегии «здорового старения». Поддержание потребления в диапазоне 334.7–420.0 мг/день представляет собой высокоэффективный, механистически обоснованный способ сохранения целостности эпителиального барьера и профилактики системных возрастных патологий.