1. Стратегическая роль межорганеллярных контактов в клеточном гомеостазе
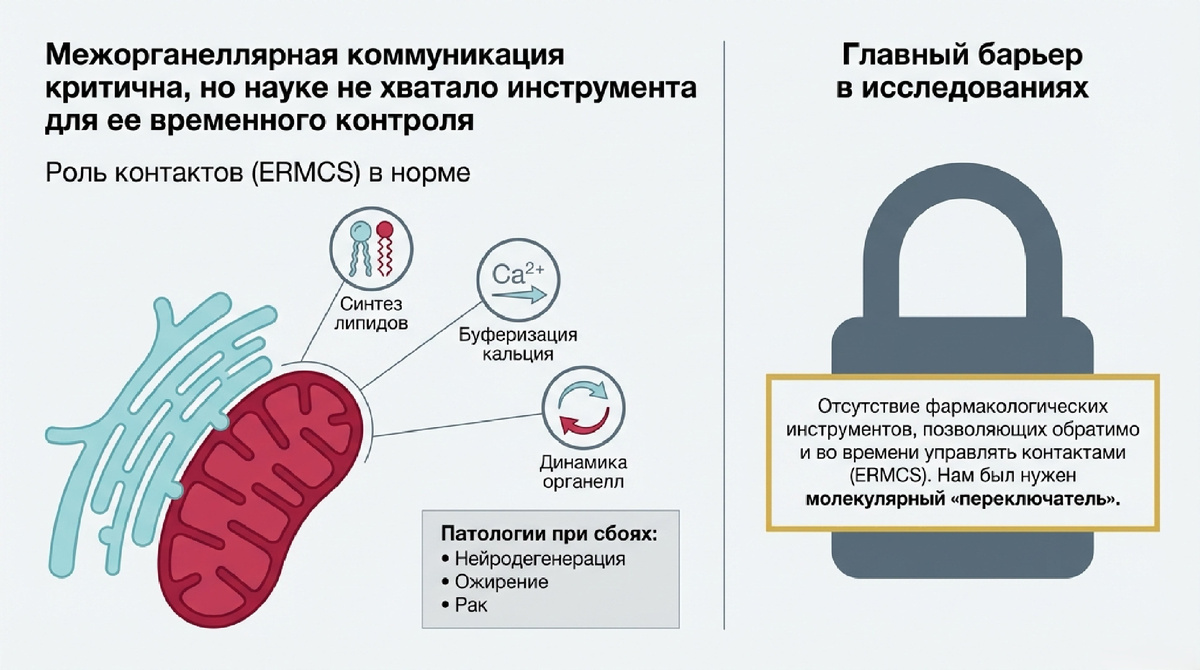
В современной молекулярной биомедицине происходит фундаментальный сдвиг парадигмы: исследовательский фокус смещается от анализа изолированных органелл к изучению динамики их взаимодействий. Ключевым узлом этой системы являются контакты между эндоплазматическим ретикулумом и митохондриями (ER-mitochondria contact sites, ERMCS). Эти структуры функционируют как высокоспециализированные метаболические платформы, координирующие транспорт липидов, кальциевую (Ca²⁺) сигнализацию и динамику митохондриальной сети.
Нарушение архитектуры ERMCS является критическим звеном в этиологии нейродегенерации, ожирения, диабета и онкогенеза. Однако до недавнего времени главным препятствием для трансляции этих знаний в клинику оставался дефицит инструментов для временного и обратимого контроля ERMCS. Обнаружение эпигенетических механизмов, управляющих физическим сближением органелл, открывает «новый рубеж» в терапии: переход от статического наблюдения к точному фармакологическому управлению межорганеллярной архитектурой.
2. Эпигенетическая ось BRD4-Mediator как главный регулятор формирования ERMCS
Исследования идентифицировали белок BRD4 (эпигенетический «ридер» ацетилированных гистонов) как центральный репрессор программы формирования ERMCS. Ингибирование BRD4 инициирует транскрипционно-зависимую перестройку клетки.
Механизм транскрипционного контроля: Фармакологическое воздействие (на примере федратиниба) вызывает вытеснение BRD4 из хроматина (в частности, из энхансерных областей PVT1, контролирующих MYC), что приводит к даунрегуляции MYC. Однако индукция ERMCS — это не побочный эффект подавления MYC, а активная программа де-репрессии, требующая участия Медиаторного комплекса. Нокдаун компонентов MED16 или MED24 блокирует формирование контактов, подтверждая, что процесс управляется скоординированным транскрипционным ответом.
Уникальный терапевтический профиль (дифференциация от стресса): В отличие от классических индукторов стресса ЭПР, механизм действия ингибиторов BRD4 (BET-ингибиторов) обладает специфическими характеристиками:
- Федратиниб (Ингибитор BRD4):Индуцирует ERMCS через Displacement-механизм (вытеснение BRD4 из хроматина).
Активирует интегрированный стресс-ответ (ISR), который является PKR-зависимым, но ISRIB-независимым.
Не активирует канонические маркеры ответа на несвернутые белки (UPR): PERK, IRE1α, ATF6 остаются неактивными.
Эффект является полностью обратимым, при этом скорость восстановления исходной архитектуры умеренно зависит от программ митофагии. - Тапсигаргин (Классический стрессор):Вызывает необратимый стресс через деплецию кальция в ЭПР.
Запускает полномасштабный UPR с активацией PERK/IRE1α.
3. Феномен «обертывания» митохондрий (ER Envelopment) и ультраструктурная перестройка
Ультраструктурный анализ (TEM, CLEM, FIB-SEM) выявил, что модуляция BRD4 приводит к формированию уникальной архитектуры — ER-mitochondria envelopment (engulfment). В отличие от стандартных точечных контактов, здесь наблюдается полное обволакивание митохондрий мембранами ЭПР.
Морфологические особенности:
- Природа мембран: В процессе участвуют преимущественно листовидные структуры шероховатого ЭПР (rough ER sheets), что подтверждается обогащением этих участков компонентами OST-комплекса (STT3A/B, RPN1).
- MOMP-подобная перестройка: Внутри «обернутых» митохондрий наблюдается коллапс крист и компартментализация внутренней мембраны (IMM), напоминающая изменения при активации апоптотической проницаемости (MOMP), но без немедленной гибели клетки.
- FAMS-опосредованная гетерогенность: Популяция митохондрий разделяется на две фракции. Примерно 15–20% митохондрий переходят в состояние SPLICSHi (полное обертывание, потеря потенциала, разрушение крист), в то время как остальные (SPLICSLo) сохраняют гомеостаз, обеспечивая выживаемость клетки.
4. Метаболическое и липидомное перепрограммирование
Физическое «обертывание» митохондрий радикально меняет их функциональный статус, создавая метаболически специализированные субпопуляции.
- Липидный парадокс: Несмотря на усиление физического контакта, наблюдается резкое истощение митохондриального фосфатидилхолина (PtdCho). Вероятно, плотное обволакивание митохондрий rough ER sheets создает стерическое препятствие для работы белков-переносчиков липидов (таких как VPS13A или STARD7), которые функционируют в условиях нормальных ERMCS.
- Протеомика SPLICSHi-митохондрий: Анализ методом FAMS выявил специфическую ферментативную сигнатуру «обернутых» органелл. Они обогащены ферментами биосинтеза пролина (PYCR1/2, ALDH18A1) и белками β-окисления жирных кислот (CPT1A, ACADVL). Это указывает на то, что индукция контактов переключает митохондрии на специфические пути катаболизма.
- Биоэнергетика: Наблюдается селективное снижение мембранного потенциала (ΔΨm) и перегрузка кальцием именно в SPLICSHi-фракции, что лимитирует адаптивную дыхательную способность клетки (SRC), создавая терапевтически значимое «окно уязвимости».
5. Роль митохондриального комплекса III и CoQ как метаболического сенсора
Способность клетки реализовать программу формирования ERMCS напрямую зависит от функционального статуса дыхательной цепи, действующей как «разрешающий фильтр».
Концепция CoQ-сенсора:
- Зависимость от ЭТЦ: Ингибирование комплекса III или IV блокирует формирование контактов. Важно отметить, что общеклеточные уровни CoQ10 при этом не меняются.
- Редокс-сигнализация: Решающим фактором является отношение CoQH₂/CoQ. Накопление восстановленной формы (убихинола) служит «сигналом запрета» для реорганизации мембран.
- Экспериментальная реверсия: Введение альтернативной оксидазы (AOX) восстанавливает способность к формированию ERMCS даже при ингибировании комплекса III, так как AOX обеспечивает альтернативный путь окисления CoQH₂.
- Локальная регуляция: Обогащение сайтов контакта ферментом COQ9 указывает на существование локальных каналов редокс-контроля, необходимых для поддержания архитектуры ER-envelopment.
6. Перспективы доклинических исследований и терапевтический потенциал
Модуляция оси «BRD4 — Mediator — Complex III» представляет собой инновационную стратегию управления клеточным метаболизмом через структурную инженерию органелл.
Стратегические преимущества:
- Прецизионность: В отличие от цитостатиков, ингибиторы BRD4 в низких дозах действуют как регуляторы архитектуры, обеспечивая временный и обратимый контроль.
- Селективная уязвимость: Создание популяции SPLICSHi-митохондрий может быть использовано для таргетного истощения метаболических резервов опухолевых клеток.
Приоритетные направления доклинических исследований:
- Ожирение и метаболический синдром: Коррекция гипертрофированных ERMCS в гепатоцитах для восстановления чувствительности к инсулину.
- Нейродегенеративные заболевания: Модуляция липидного обмена и кальциевого гомеостаза в моделях болезней, связанных с патологией контактов.
- Прецизионная онкология: Использование «липидного парадокса» и метаболического перепрограммирования для синергии с существующими схемами химиотерапии.
Заключение: Выявление транскрипционно-метаболического реостата, управляющего контактами ERMCS, переводит управление клеточной архитектурой в разряд доступных фармакологических задач. Использование BET-ингибиторов как инструментов модуляции межорганеллярной коммуникации открывает новую главу в системном восстановлении метаболического здоровья.