1. Введение: Эволюционная роль митохондрий в иммунном ответе
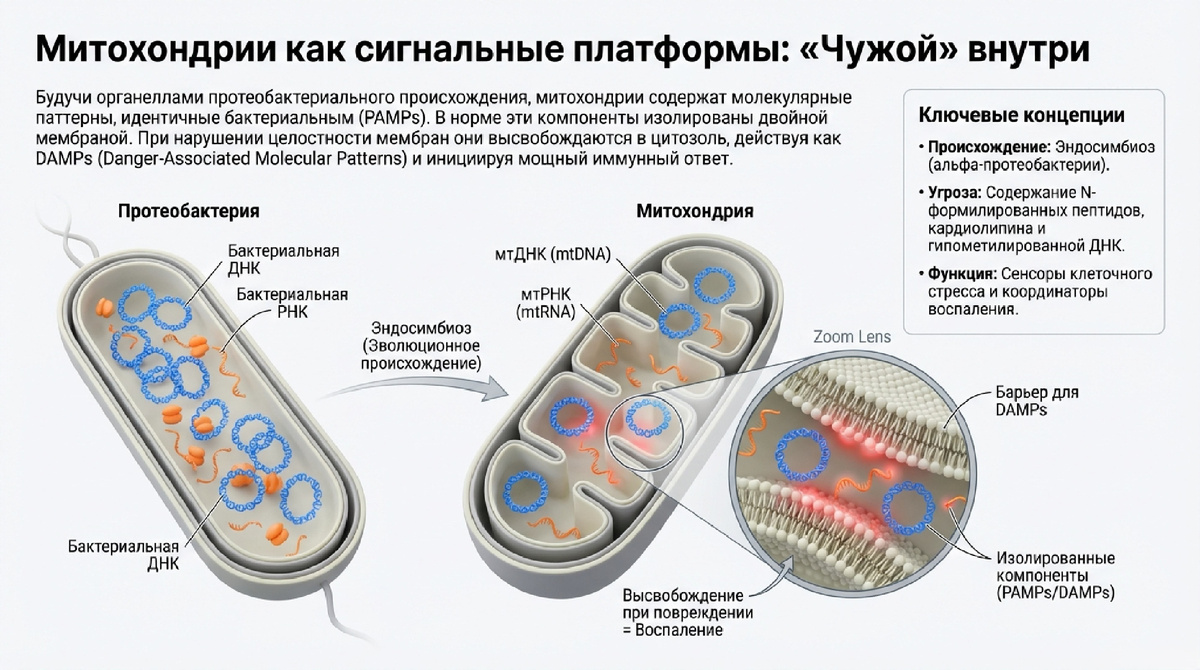
В современной парадигме молекулярной иммунологии митохондрии рассматриваются не просто как метаболические узлы, а как центральные «сигнальные органеллы», интегрирующие гомеостатические стимулы и стрессовые сигналы. Стратегическая роль митохондрий в иммунитете неразрывно связана с их эндосимбиотическим происхождением. Будучи потомками древних протеобактерий, эти органеллы сохранили специфические прокариотические черты (бактериальное наследие), которые врожденная иммунная система эукариот идентифицирует как признаки инвазии или критического повреждения.
Обладая высокой структурной пластичностью, митохондрии постоянно меняют свою морфологию через циклы деления и слияния, формируют динамичные контакты с эндоплазматическим ретикулумом (MERCS) и перемещаются по цитоскелету. Эта мобильность позволяет органеллам выступать в качестве сенсоров, которые не только фиксируют локальный стресс, но и транслируют его на системный уровень. При нарушении целостности митохондриальных мембран происходит высвобождение молекул, которые в норме изолированы в матриксе или межмембранном пространстве (IMS). Эти молекулы, действуя как митохондриальные DAMPs (danger-associated molecular patterns), становятся фундаментальными триггерами воспалительного ответа.
2. Репертуар митохондриальных паттернов (DAMPs)
Выход митохондриальных компонентов в цитозоль или внеклеточную среду интерпретируется клеткой как сигнал опасности именно из-за их структурного сходства с патоген-ассоциированными молекулярными паттернами (PAMPs). Специфический иммунный ландшафт формируется набором следующих мессенджеров:
• мтДНК (mtDNA): Кольцевая молекула, содержащая гипометилированные CpG-мотивы. Из-за отсутствия гистоновой защиты и близости к источнику активных форм кислорода (АФК) в дыхательной цепи, мтДНК крайне подвержена окислению. Особый интерес представляет переход B-формы мтДНК в Z-форму (Z-mtDNA) при торсионном стрессе, что критично для специфической рецепции.
• мтРНК (mtRNA): Продукт двунаправленной транскрипции митохондриального генома. При нарушении деградации РНК-деградосомой формируются иммуностимулирующие двуцепочечные структуры (dsRNA). Важнейшим дифференциатором здесь является механизм экспорта: за эффлюкс mt-dsRNA в цитозоль отвечает транслокатор адениновых нуклеотидов-2 (ANT2), функционирующий как двунаправленный РНК-транслокон в составе внутренней мембраны (IMM).
• Кардиолипин: Уникальный фосфолипид IMM. Его транслокация на внешнюю мембрану (OMM) или выход во внеклеточную среду служит сигналом для митофагии или сборки инфламмасом. Кардиолипин проявляет функциональную вариативность в зависимости от степени окисления и состава ацильных цепей.
• N-формилированные пептиды: Продукты митохондриальной трансляции, сохраняющие N-формилметионин, что делает их структурными аналогами бактериальных белков и позволяет напрямую активировать рецепторы формилпептидов.
• Митокины (Mitokines): Пептиды, такие как MOTS-c и гуманин, кодируемые короткими открытыми рамками считывания в мтДНК, участвующие в ретроградном сигналинге и адаптации к метаболическому стрессу.
Эффективность инициации ответа зависит от конкретного пути эвакуации этих DAMPs, который определяется характером и интенсивностью клеточного повреждения.
3. Архитектура транслокации: пути выхода DAMPs в цитозоль
Преодоление двухмембранного барьера митохондрий является жестко регулируемым процессом. В зависимости от типа стресса клетка использует различные механизмы — от формирования селективных пор до крупномасштабной герниации мембран.
Тип канала/поры / Механизм и специфика / Пропускаемый субстрат
Мегапоры BAX/BAK / Олигомеризация при подавлении каспаз; расширение пор ведет к герниации IMM в цитозоль (механизм CICD). / Цельные нуклеоиды мтДНК, mtRNA, цитохром c, SMAC.
Ось VDAC/mPTP / Координированное открытие поры mPTP (IMM) и пор VDAC (OMM). Регулируется гексокиназой и белками семейства Prohibitin. / Фрагментированная и окисленная мтДНК.
Поры Газдерминов (GSDMD/E) / N-терминальные фрагменты GSDMD (активированные каспазой-1) или GSDME (каспазами-3/7) встраиваются в OMM/IMM. / Небольшие фрагменты мтДНК (поры слишком малы для цельных нуклеоидов), mtRNA.
Митохондриальные везикулы (MDVs/VDIMs) / Отпочковывание везикул (в т.ч. ESCRT-зависимых VDIMs) для транспорта в лизосомы или внеклеточную среду. / Селективно упакованные белки и фрагменты мтДНК.
Эти механизмы определяют, какой именно рецептор врожденного иммунитета будет активирован в цитоплазме, запуская специфическую программу ответа.
4. Рецепция и внутриклеточные сигнальные каскады
Точное распознавание митохондриальных лигандов паттерн-распознающими рецепторами (PRRs) трансформирует физическое повреждение органеллы в биохимический каскад.
1. Путь cGAS/STING и роль ZBP1: мтДНК связывается с cGAS, стимулируя синтез cGAMP, который активирует STING на мембране ЭПР. После транслокации в Гольджи STING рекрутирует TBK1 для фосфорилирования IRF3. Важным дополнением является сенсор ZBP1, который специфически детектирует Z-mtDNA, возникающую при стрессе, и совместно с cGAS усиливает продукцию интерферонов I типа через взаимодействие с RIPK1/RIPK3.
2. Инфламмасома NLRP3: Окисленная мтДНК и кардиолипин способствуют сборке NLRP3. Процесс сопряжен с ремоделированием крист и закрытием их соединений, что блокирует выход цитотоксичного цитохрома c, переключая клетку с апоптоза на воспалительный пироптоз.
3. Сигналинг RLR/MAVS: mt-dsRNA распознается рецепторами MDA5/RIG-I, которые взаимодействуют с белком MAVS, заякоренным на OMM. Митохондрия здесь служит платформой для сборки «сигналосомы», активирующей как IRF3/7, так и NF-κB.
4. Митохондриальная платформа NF-κB: Митохондрии выступают в роли «транспортных средств» для активированного NF-κB. При активации рецепторов TNFR1 на OMM собирается комплекс, включающий LUBAC, NEMO и PINK1. Стабилизированный PINK1 фосфорилирует убиквитиновые цепи на NEMO, предотвращая их гидролиз деубиквитиназой OTULIN. Этот комплекс физически перемещается к ядру по микротрубочкам, что критически важно в поляризованных клетках (нейронах).
5. Эндосомальный TLR9: Гипометилированная мтДНК, доставленная через MDVs или эндоцитоз, распознается TLR9, активируя митоген-активируемые протеинкиназы (MAPK) и NF-κB.
5. Патофизиологическое значение и «хронизация» воспаления
Длительная экспозиция митохондриальных DAMPs трансформирует адаптивный ответ в драйвер системных патологий.
1. Старение и SASP: Снижение эффективности митофагии ведет к «утечке» мтДНК/мтРНК. Это поддерживает провоспалительный секреторный фенотип (SASP), обеспечивая состояние стерильного воспаления (inflammaging).
2. Аутоиммунные заболевания: При системной красной волчанке окисленная мтДНК напрямую связывает GSDMD, усиливая его олигомеризацию и выброс нейтрофильных экстраклеточных ловушек (NETs), что замыкает порочный круг аутовоспаления.
3. Противоопухолевый иммунитет и «Minority MOMP»: В условиях умеренного стресса может возникать «миноритарный MOMP» (сублетальная пермеабилизация мембран), не приводящая к смерти клетки. Подавление эффекторных каспаз (3 и 7) предотвращает деградацию DAMPs, позволяя BAX/BAK-порам расширяться. Это превращает потенциально «тихий» апоптоз в иммуногенную клеточную смерть, активируя MDA5-зависимый интерфероновый ответ, стимулирующий CD8+ Т-клетки против опухоли.
6. Заключение: Перспективы терапевтического воздействия
Митохондриальный сигналинг — это динамическая система, обеспечивающая баланс между выживанием и воспалением. Понимание механизмов этого баланса открывает возможности для прецизионной терапии.
Перспективные направления исследований включают:
• Мультимодальное управление DELE1: Полноразмерный L-DELE1 на внешней мембране выступает сенсором стресса импорта белков, тогда как расщепленный протеазой OMA1 фрагмент S-DELE1 (олигомеризующийся в октамеры) транслирует стресс матрикса. Модуляция этой оси через киназу HRI позволит управлять интегрированным стресс-ответом (ISR).
• Таргетирование пор VDAC: Блокада селективного выхода фрагментированной мтДНК может стать стратегией борьбы с хроническим воспалением при нейродегенерации (например, при БАС, где TDP-43 провоцирует выход мтДНК).
• Модуляция STING и ZBP1: Ингибирование этих сенсоров необходимо для купирования интерферонопатий и миокардиальной токсичности, вызванной химиотерапией.
Дальнейшее изучение митохондрий как «сигнальных хабов» позволит перевести фундаментальные знания о врожденном иммунитете в плоскость эффективного клинического контроля гомеостаза.