Предисловие. Я пытался несколько раз опубликовать эту статью в течение нескольких лет, однако получал отказы от Журналов. В статье, безусловно, есть свои слабости - скудные материалы, не позволяющие сделать полноценную статистическую обработку и полноценные выводы.
В то же время, статья является уникальной по своей сути, поскольку мне удалось сделать основательный обзор литературы и убедиться в верном движении моей научной мысли.
В августе 2025 года я ездил на место своих полевых работ и увидел, как сбылись мои прогнозы описанные в диссертации, которую защитил в 2022 году. Об этом расскажу позднее.
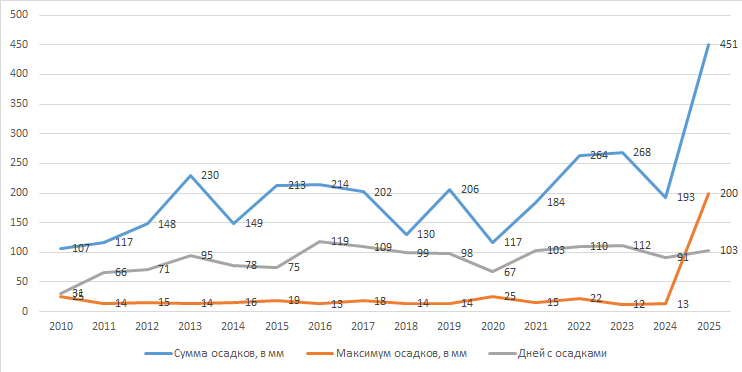
Изначально я не планировал ничего публиковать, но важным аргументом для изменения решения стали климатические показатели 2025 года, которые проанализировал в начале 2026 года. Данные оказались уникальными, т.к. за 12 месяцев выпал 451 мм осадков. Для Астраханской области это огромные цифры.
Далее я также продолжу наблюдать за тем, что происходит на территории Астраханской области и делать публикации на этот счёт.
Ниже статья, что не опубликована.

ВЛИЯНИЕ ПАРАЗИТОВ НА РАЗНОЦВЕТНУЮ ЯЩУРКУ
EREMIAS ARGUTA DESERTI (GMELIN, 1789)
В ЮГО-ВОСТОЧНОЙ ЧАСТИ НИЖНЕГО ПОВОЛЖЬЯ
Аннотация. При исследовании весной 2017 года разноцветной ящурки Eremias arguta deserti (Gmelin, 1789) у особей обнаружены эндо и эктопаразиты. Статистический анализ подтвердил наличие у разноцветной ящурки групп как среди самцов, так и среди самок с эктопаразитами и без. Клещи и черви найдены у особей обоих полов в возрасте не менее 2 лет, в то время как у неполовозрелых животных они не выявлены. Также весной 2017 года найдены следы от укусов на теле самцов. В иные периоды исследований укусы на теле самцов не фиксировались. Кроме того этот полевой сезон E. arguta deserti имела достоверные группы самцов и самок с укусами на теле и без. Укусы и травмы на телах самок в последующие брачные сезоны существенно снизились. Единичные за весь период полевых работ наблюдения и результаты на фоне паразитизма и не свойственной для исследуемой группы агрессивности дают повод предположить о наличии взаимосвязи между паразитизмом, агрессивностью и результатами сезона размножения разноцветной ящурки весной 2017 года. Данный вопрос требуется дополнительного изучения.
В поселении круглоголовки-вертихвостки Phrynocephalus guttatus guttatus (Gmelin, 1789), обитавшей на совместной с разноцветной ящуркой территории, паразиты не обнаружены. Наблюдения в брачный сезон 2017 года свидетельствуют о присутствии в аридной экосистеме стенобионтных паразитов.
Ключевые слова: паразит, агрессивность, популяция, разноцветная ящурка, Eremias arguta deserti, круглоголовка-вертихвостка, Phrynocephalus guttatus guttatus, Нижнее Поволжье.
Введение
Среди факторов окружающей среды биотическая связь «хозяин-паразит» не редко играет решающую в роль в жизненном цикле особи и популяции. Паразит использует тело хозяина как среду обитания и (или) в качестве пищи. В природе число хозяев для паразитов вполне ограничено, поэтому биологические, физические и химические показатели окружающей среды, а также экология потенциального хозяина имеют большое значение [13]. Известны исследования, описывающие положительное влияние количества осадков и температуры на гельминтов, а также промежуточных хозяев [31, 35].
Климатические изменения, фиксируемые в различных точках нашей планеты и их последствия, приводят в напряжение все доступные иммунные и адаптационные механизмы живого вещества [6], меняют потоки вещества и энергии в экосистемах. С точки зрения прикладной классификации экологических факторов А.С. Мончадского [18] паразитизм относится к изменяющимся без закономерной периодичности факторам и как правило, случаен.
У самцов позвоночных животных чаще, чем у самок отмечаются паразитарные заболевания [37]. Ряд учёных сравнивает паразитов у ящериц одного вида в различных географических точках [34], отмечает более высокое поражение гельминтами [40, 41] и клещами [14] самцов, однако есть данные о равном распределении нагрузки паразитов между полами [30]. Исследования в разные времена года показали неодинаковое распространение паразитов среди полов. Так, в июле-августе, у самок находили больше нематод, в апреле у самцов [37]. При исследовании ящериц Podarcis bocagei и Podarcis carbonelli, также найдена взаимосвязь между сезонами и численностью гельминтов [32].
При заражении гекконов Quedenfeldtia trachyblepharus (Boettger, 1874) клещами установлена зависимость от сезона наблюдений, высотой над уровнем моря, полом и размером хозяина [38]. Здесь же выявлено поражение у 35% молодняка одним видом паразитов, в то время как у взрослых обнаружено до трёх видов паразитов.
Подчёркивается повышенная паразитическая нагрузка на взрослых особей, в отличие от молодняка [32], поскольку они имеют большую площадь тела и дают большую среду обитания для паразитов [50]. Роль размеров тела отмечена и другими исследователями [35]. Стоит отметить, что у другого вида ящериц не выявили корреляцию между числом паразитов и полом, возрастом и даже размером тела [33].
Высокая роль отводится рациону питания, как основному источнику паразитов. В литературе имеется обоснование разнообразия гельминтофауны в зависимости от травоядного или плотоядного типа питания пресмыкающихся, указывается на меньшее видовое разнообразие паразитов у плотоядных рептилий [48]. Между тем, результаты исследований более 350 видов ящериц из 30 семейств дало обратный результат и дополнительно выявило меньшее число видов гельминтов у травоядных ящериц (особенно в личиночной стадии) [43].
Также описывают более высокую восприимчивость партеногенетических видов к паразитам, чем у их симпатрических половых родственников, [46]. Однако другие авторы считают необходимым провести больше исследований по данному вопросу [49].
Есть мнение о преобладании нематод среди хозяев-лацертид, а само биоразнообразие паразитофауны ограничено пойкилотермностью рептилий [36]. Преобладание нематод у ящериц описано в работах некоторых исследователей [47, 33] и в частности это отмечено у разноцветной ящурки Eremias arguta (Pallas, 1773) [29, 8].
Разноцветная ящурка является плотоядным животным, питающимся насекомыми [11, 10, 26]. В монографии Н.Н. Щербака с соавторами [29] есть сведения о растительных (молочай, полынь и др.) и неорганических остатках (песчинки, кусочки глины и др.), найденных в желудках ящериц и попавших туда случайно. Отмечены случаи каннибализма.
Первым фундаментальным трудом по обобщению данных о гельминтах у пресмыкающихся, обитающих на территории СССР, является монография В.П. Шарпило [27]. Автор описал у разноцветной ящурки E. arguta 18 видов червей.
В труде Н.Н. Щербака с соавторами [29] описано 8 видов клещей и 19 видов гельминтов. З.К. Брушко [4] несколько расширила сведения о паразитах E. arguta. С.В. Ганщук [8], проводившая исследования гельминтофауны пресмыкающихся на территории Астраханской области в апреле-мае 2012 года, обнаружила у E. arguta 5 видов гельминтов. В материалах по Азербайджану (окрестности г. Ширван) [7] у разноцветной ящурки найдены ооцисты – C. аff. serpentis.
В Ленкоранской природной области юго-восточного Азербайджана при исследовании полосатой ящерицы найдены гельминты, для которых разноцветная ящурка является хозяином [19].
В проведённых ранее многолетних исследованиях удалось показать каким образом преимущественные [12] климатические показатели, в первую очередь осадки, повлияли на экосистему аридной территории юго-восточной части Нижнего Поволжья, и как ящерицы отреагировали на происходившие изменения [15].
Целью данной работы стала попытка описать с точки зрения биоэкологии изменения во внутрипопуляционной группировке разноцветной ящурки E. arguta deserti на фоне ярко выраженного паразитизма весной 2017 года в сопоставлении с данными, полученными в другие периоды полевых работ, когда паразитизм не выявлялся, а также сравнить с жизнедеятельностью особей круглоголовки-вертихвостки Ph. guttatus guttatus, обитавших на той же территории.
Материалы и методы
Район исследований расположен вблизи координат: N 46.913954, E 47.932622 и представляет собой северный подтип пустынного типа ландшафта с бугристо-грядовыми и бугристо-грядово-барханными песками [1]. Основными эдификаторами [5] в этот период экологических изысканий являлись джузгун безлистный (Calligonum aphyllum Litv.) и полынь песчаная (Artemisia arenaria DC.). В 2018 и 2019 году видовой состав растений обогатился новыми видами, и фитоценоз представлял собой уже джузгунно-разнотравное сообщество с явными признаками сукцессии в сторону степного фитоценоза [17].
В работе использованы материалы, полученные по результатам наблюдений в период: с 28.08.2016 по 12.09.2016, с 07.05.2017 по 26.05.2017, с 28.08.2017 по 13.09.2017, с 06.05.2018 по 18.05.2018, с 19.08.2018 по 29.08.2018, с 04.05.2019 по 15.05.2019.
Помимо меруса [25] разноцветной ящурки E. arguta deserti, на территории исследования обитала также и внутрипопуляционная группировка [28] номинативного подвида круглоголовки-вертихвостки Ph. guttatus guttatus. Ранее здесь встречали и ушастую круглоголовку Phrynocephalus mystaceus mystaceus (Pallas, 1776), однако ко времени проведения исследований она вымерла [15].
Все отловленные ящерицы маркировались временной и постоянной меткой. Рептилиям определяли пол, присваивали уникальный номер, который наносили на спину спиртовым маркером чёрного цвета. Поскольку животные несколько раз за год сбрасывали наружный роговой слой и временная метка исчезала, использовали постоянное мечение путём отрезания кончиков пальцев [45, 51]. Наблюдения за особями не выявили существенного влияния на дальнейшую активность и жизнь пресмыкающихся.
Места отлова и последующих встреч с животными наносили на карту. Особь, встреченную более двух раз в пределах одной территории, считали оседлой, в то время как животное, встреченное только один раз, относили к мигрантам. В случае же, если животное встречалось только два раза, за ним проводили дополнительные наблюдения, чтобы оценить степень его знакомства с биотопом: наличие убежищ, перемещение по территории, контакты с окружающими объектами [15, 21]. Данная методика успешно применялась на протяжении нескольких лет и показала свою эффективность.
Особей также взвешивали с точностью до 0,1 г, измеряли длину тела: от кончика морды до клоаки и от клоаки до кончика хвоста с точностью до 1 мм.
Определение возраста животных осуществлялось на основе трудов А.М. Сергеева [22, 23, 24] по постэмбриональному росту рептилий, личных полевых наблюдений [15] и результатов других авторов [2, 3, 20].
Поскольку исследование паразитофауны не являлось основной целью полевых работ, то в нашем распоряжении не оказалось нужного инвентаря для сбора и хранения образцов. Таким образом, виды паразитов не установлены. Между тем, нам удалось установить всех особей с эктопаразитами, в то время как особей с эндопаразитами фиксировали только после обнаружения червей в экскрементах или при визуальном осмотре клоаки. Стоит обратить внимание – за все предыдущие и последующие периоды исследований у разноцветной ящурки и круглоголовки-вертихвостки паразиты не зафиксированы.
Общее число промаркированных E. arguta deserti составило весной 2017 года – 76 особей, весной 2018 года – 38 особей, весной 2019 года – 12 особей, осенью 2017 года – 25 особей, осенью 2018 года – 14 особей. Полученные результаты систематизировали и обрабатывали с помощью программного обеспечения Microsoft Office Excel.
Статистическая обработка данных проведена с помощью критерия Хи-квадрат в программе STATISTICA 12.
Результаты и их обсуждение
Как известно, разноцветная ящурка при преследовании нередко выделяет экскременты и пытается укусить преследователя [29]. Только весной 2017 года установлен гельминтоз у восьми ящурок (до 15 гельминтов в экскрементах самца и до 10 гельминтов в экскрементах самки), из них 5 оседлых (♀ – 2; ♂ – 3) и 3 мигранта (♀ – 1; ♂ – 2). Найдены клещи, как правило, в области брюха (до 22 клещей на теле самца и до 2 клещей на теле самки) у 15 ящурок, из них 6 оседлых (♀ – 1; ♂ – 5) и 9 мигрантов (♀ – 4; ♂ – 5). Только у одного оседлого самца одновременно зафиксированы клещи (1 экземпляр) и гельминтоз (7 гельминтов в экскрементах) (рис. 1).
Разница внутри группы самок E. arguta deserti с эктопаразитами и без статистически достоверна (p<0,05). Аналогичная картина у самцов. При сравнении всей группировки животных обоих полов с наличием клещей и их отсутствием результаты оказались не достоверными (0,06).
При анализе статистической значимости у оседлых и мигрирующих животных с наличием (отсутствием) эктопаразитов выявлено, что из числа мигрирующих особей меруса разноцветной ящурки имеется статистическая значимость внутри группы самцов и внутри группы самок. Однако результаты внутри всей группы особей как самцов, так и самок оказались не достоверными.
Среди оседлых особей картина значительно отличается от данных, полученных по мигрантам. Так, среди всех оседлых особей, как самцов, так и самок наличие групп животных с клещами и без оказалось достоверным. Такие же сведения получены по самцам. У самок результаты оказались статистически не значимы.
Провести аналогичные исследования по эндопаразитам в данном исследовании невозможно из-за не большой выборки, однако роль гельминтов в разрезе подобных явлений ещё предстоит определить дополнительными работами.
Рис. 1. Клещ на теле (a) и гельминты в экскрементах (b) у E. argita deserti весной 2017 года
Минимальная длина туловища особи, заражённой гельминтами, составила 65 мм (самец второго года жизни), максимальная – 71 мм (самка третьего года жизни). Минимальная длина туловища особи, заражённой клещами, составила 55 мм (самка второго года жизни), максимальная – 72 мм (самец третьего года жизни). У полувзрослых особей паразиты не обнаружены.
Подобного паразитизма, как у разноцветной ящурки весной 2017 года, у круглоголовки-вертихвостки не выявлено ни в один из исследуемых сезонов. Численность Ph. guttatus guttatus весной 2017 года составила – 44 особи, осенью 2017 года – 53 особи, весной 2018 года – 38 особей, осенью 2018 года – 35 особей, весной 2019 года – 21 особь.
В рамках полевых работ отловлены и промаркированы две круглоголовки-вертихвостки, ушедшие на расстояние 135 метров (осенью 2016 года) и 100 метров (весной 2017 года) от первичного места учёта [15].
Так, осенью 2016 года самка впервые отловлена 30.08.2016 в 13:04, повторно 07.09.2016 в 10:31 по местному времени. Самец был впервые отловлен в тот же сезон, что и самка 29.08.2016 в 11:57, повторно 10.05.2017 в 07:56 по местному времени. Самка двигалась строго на юго-восток, самец на восток-северо-восток.
Наблюдения за перемещениями данных особей подтверждают использование экологических коридоров, переходных зон (экотонов), между стациями [9] других животных и в том числе, разноцветной ящурки. Соответственно круглоголовка-вертихвостка имела возможность стать объектом для эндо и эктопаразитов, как и разноцветная ящурка.
По нашим данным [16] агрессивность у животных отмечена только между самцами Ph. guttatus guttatus, в то время как в группировке E. argura deserti какие-либо столкновения отсутствовали. За весь период полевых исследований зафиксирована единичная демонстративная попытка Ph. guttatus guttatus атаковать E. argura deserti при попытке последней особи пробежать по открытой территории участка первой.
Таким образом, межвидовая конкуренция объективно не проявлялась, поскольку является общеизвестным факт о разных стациях круглоголовки-вертихвостки и разноцветной ящурки.
Полученные результаты по миграции круглоголовки-вертихвостки и об особенностях проявления паразитизма позволяют вновь сказать о различии в пищевых нишах [16]. Поскольку при исследовании гельминтов Gallotia atlantica (Peters et Doria, 1882) с островов Лансароте и Фуэртевентура на Канарском архипелаге, Испания, выявлены универсальные паразиты (убиквисты, эврибионты) способные жить за счёт хозяев из разных семейств рептилий [44], то зафиксированные в момент исследований паразиты у разноцветной ящурки можно отнести к узкоспециализированным.
В течение всех периодов наблюдений, общая численность E. arguta deserti постепенно сокращалась (табл. 1) [15].
Весной 2017 года из 34 самцов 2 оказались полувзрослыми, а остальные двухлетками. У самок картина получилась следующая: 6 полувзрослых ящериц и 36 двухлеток. Весной 2018 года полувзрослыми оказались только 3 самки. Среди двухлеток оказалось 13 самцов и 22 самки. Весной 2019 года полувзрослые ящерицы не найдены.
Численность сеголеток осенью 2017 года (первая группа сеголеток насчитывала 8 особей, вторая – 6) несильно отличалась от численности сеголеток осени 2018 года (первая группа сеголеток насчитывала 1 ящерицу, вторая – 9), несмотря на существенную разницу в численности животных, самок в частности, в сезоны размножения 2017 и 2018 гг [15].
Исходя из данных полученных по живородящей ящерице Zootoca vivipara, агрессивность самцов вызывала стресс у самок, снижала их плодовитость и выживаемость [42]. В то же время, у ящерицы Tropidurus hispidus не выявили зависимость между плодовитостью, энергетическими затратами и паразитами [40].
Влияние нематод на поведение выявлено у ящериц Egernia stokesii. Животные, которым давали антигельминтовые препараты, выздоравливали, дольше передвигались, а также чаще уделяли время баскингу [39]. Больные животные больше времени проводили в убежищах и меньше грелись на солнце.
По результатам анализа данных весны 2017 года с помощью критерия Хи-квадрат статистическая значимость подтвердилась (p<0,05):
- между группой покусанных самок и группой покусанных самцов;
- между группой покусанных самок и группой самок без следов укуса на теле (также выявлено и весной 2018 года);
- между группой покусанных самцов и группой самцов без следов укуса на теле.
Таким образом, мы предполагаем, что избыточная агрессивность самцов, вызванная экто и эндо паразитами в весенний сезон 2017 года, оказала влияние на самок и снизила результаты сезона размножения.
Так, только весной 2017 года обнаружены укусы на телах самцов: 4 самца имели ссадины только на хвосте и у основания хвоста, 1 самец имел ссадины на голове и хвосте, 1 самец имел ссадины на голове, шее и хвосте, 1 самец имел ссадины только на голове и 1 самец имел ссадины только в области головы и брюха.
В то же время у самок, во все брачные периоды, обнаружены следы укусов на теле. Стоит подчеркнуть, что весной 2017 года только одна полувзрослая самка имела укусы на хвосте и у основания хвоста, у одной двухлетней особи найдены укусы в области брюха. Весной 2018 года у полувзрослых особей укусы на теле не найдены. В целом, весной 2017, 2018 и 2019 года укусы у самок обнаружены, в основном, на хвосте и у основания хвоста (табл. 2).
Травмы у самцов и самок, с точки зрения места их нанесения, отличаются. На телах самцов преобладали травмы на голове и шеи, укусы на хвосте и у основания хвоста. У самок, практически все травмы, зафиксированы на хвосте и у основания хвоста. Здесь вызывает интерес характер травм у самцов и самок весной 2017 года, а также высокий уровень агрессии самцов друг к другу (преимущественно у мигрантов). Ссадины (на голове, шее, хвосте и у основания хвоста), возможно, говорят об атаке самцов друг на друга не просто чтобы прогнать конкурента, а для спаривания. Однако, агрессор отпускал конкурента после получения обратной реакции об ошибочности выбора полового партнёра.
Здесь также следует сказать и о поведении E. arguta deserti во время отлова и маркировки особей. Ящерицы, особенно самцы, ожесточённо сопротивлялись, кусали пальцы до крови и яростно вырывались из рук. Именно такое несвойственное для данного поселения поведение разноцветной ящурки повторно заставило обратить внимание на невысокую разницу в численности сеголеток осенью 2017 года (14 сеголеток при общей численности самок в 42 особи) и осенью 2018 года (10 сеголеток, при общей численности самок в 25 особей) (табл. 1).
Очевидно сыграли роль не только лишь укусы самцов, так как сжатие самцами самок челюстями в области основания хвоста является обычным процессом при спаривании. Здесь стоит сделать упор на силу и характер укусов. К сожалению, число укусов на телах самок не считали. Сила, с которой самцы и самки могут сжимать челюсти также не известна, однако самки, подвергавшиеся укусам со стороны самцов, не редко имели дряблую, жёваную кожу у основания хвоста. Подобной деформации эпителий у самок в другие сезоны не отмечено.
Заключение
1. Весной 2017 года достоверно выявлены группы особей E. arguta deserti с эктопаразитами и без. Обнаружены особи и с эндопаразитами, однако из-за небольшой выборки дать статистическую оценку не представляется возможным.
2. У мигрирующей части меруса разноцветной ящурки с наличием (отсутствием) клещей на теле выделены достоверные группы как среди самцов, так и среди самок. У оседлых особей данные внутри группы самок не достоверны. Однако статистически значимы оказалось наличие групп с клещами и без при сравнении данных самок с данными самцов, а также самцов с самцами. У полувзрослых особей паразитизм не выявлен.
3. Результаты, полученные на основании анализа укусов на телах разноцветной ящурки весной 2017 года, оказались достоверными у всех сравниваемых групп: самки-самцы, самки-самки, самцы-самцы. Весной 2018 году достоверными стали результаты только среди группы самок.
4. Следует предположить, что влияние паразитов привело к повышенной агрессивности в поселении разноцветной ящурки и снизило результаты сезона размножения весны 2017 года, как это выявляли и у других видов. Описанные результаты требуют дополнительных исследований.
5. У круглоголовки-вертихвостки, несмотря на возможность миграции через стации разноцветной ящурки, паразитизм не выявлен. Таким образом на территории находились паразиты-стенобионты.
Литература
1. Атлас Астраханской области. М.: Роскартография, 1997. 48 с.
2. Бадмаева В.И. Ящерицы Калмыкии : Дис. … канд. биол. наук. Киев, 1983. 227 с.
3. Банников А.Г., Даревский И.С., Ищенко В.Г., Рустамов А.К., Щербак Н.Н. Определитель земноводных и пресмыкающихся фауны СССР. М.: Просвещение, 1977. 415 с.
4. Брушко З.К. Ящерицы пустынь Казахстана. Алматы: Конжык, 1995. 231 с.
5. Быков Б.А. Геоботанический словарь // Алма-Ата: Наука, 1973. 215 с.
6. Вернадский В.И. Живое вещество. М.: «Наука», 1978. 358 с.
7. Гаибова Г.Д., Мамедова С.О. Кишечная кокцидиофауна (Apicomplexa: Coccidia) рептилий Азербайджана и ее формирование под воздействием антропогенных факторов // Амурский зоологический журнал. 2021. Т. XIII, Вып. 3. С. 353–368. https://doi.org/10.33910/2686-9519-2021-13-3-353-368.
8. Ганщук С.В. Гельминтофауна пресмыкающихся Северного Прикаспия // Вестник ТГУ. 2013. Т. 18, Вып. 6. С. 2992–2993.
9. Гептнер В.Г. Общая зоогеография. М.: Биомедгиз. 1936. 553 с.
10. Гончаров А.Г. Характеристика питания разноцветной ящурки Eremias arguta (Pallas, 1773) на севере ареала // Вестник ТГУ. 2013. Т. 18, Вып. 6. С. 2994–2996.
11. Дебело П.В., Чибилёв А.А. Амфибии и рептилии Урало-Каспийского региона // Серия: Природное разнообразие Урало-Каспийского региона. Т.III. Екатеринбург: РИО УрО РАН, 2013. 400 с.
12. Кашкаров Д.Н. Среда и сообщество (основы синэкологии). М.: Государственное медицинское издательство, 1933. 242 с.
13. Кеннеди К. Экологическая паразитология. М.: изд-во «Мир», 1978. 231 с.
14. Кидов А.А., Тимошина А.Л., Матушкина К.А., Коврина Е.Г. Паразитизм европейского лесного клеща Ixodes ricinus (Linnaeus, 1758) (Acari, Parasitiformes: Ixodidae) на ящерице Браунера, Darevskia brauneri (Mehely, 1909) (Reptilia, Sauria: Lacertidae) // Вестник Бурятского государственного университета. Биология. География. 2013. Вып. 4. С. 165–166.
15. Мишустин С.С. Динамика структуры популяций ящериц в полупустынях юго-восточной части Нижнего Поволжья // Дис. … канд. биол. наук. Москва, 2022. 180 с.
16. Мишустин С.С. Этологическая структура группировок круглоголовки-вертихвостки и разноцветной ящурки в период размножения при совместном обитании // Материалы Международного молодёжного научного форума «ЛОМОНОСОВ-2019». 2019. С. 1.
17. Мишустин С.С., Полынова Г.В. Динамика фитоценозов в полупустынях юго-восточной части Нижнего Поволжья // Вестник института ИКИАТ. 2019. Т. 1, Вып. 38. С. 10–11. https://doi.org/10.24411/2071-7830-2019-10002.
18. Мончадский А.С. Экологические факторы и принципы их классификации // Журн. общей биологии. 1962. Т. 23, Вып. 5. С. 370–380.
19. Мусаева С.М. Гельминтофауна полосатой ящерицы в Ленкоранской природной области и ландшафтно-экологический анализ её распространения // Юг России: экология, развитие. 2012. Т. 7, Вып. 3. С. 48–53. https://doi.org/10.18470/1992-1098-2012-3-48-53.
20. Окулова Н.М. Некоторые черты биологии разноцветной ящурки в Западном Казахстане // Вопросы герпетологии. Л.: Изд-во Ленингр. Ун-та, 1964. С. 50–51
21. Полынова Г.В., Бажинова А.В., Гриб Е.В. Материалы по пространственной структуре популяции круглоголовки-вертихвостки Phrynocephalus guttatus guttatus в полупустынях Астраханской области // Материалы международной конференции, посвящённой памяти Н.А. Зарудного. 2012. С. 260–264.
22. Сергеев А.М. Материалы к вопросу о постэмбриональном росте рептилий // Зоол. журн. 1937. Т. XVI, Вып. 4. С. 723–735.
23. Сергеев А.М. Материалы по постэмбриональному росту рептилий // Зоол. журн. 1939. Т. 18, Вып. 5. С. 888–903.
24. Сергеев А.М. Эволюция эмбриональных приспособлений рептилий. –М.: Советская наука, 1943. 203 с.
25. Флинт В.Е. Пространственная структура популяций мелких млекопитающих. М.: Наука, 1977. 181 с.
26. Чирикова М.А., Казенас В.Л. Амфибии и Рептилии. // Серия «Животные Казахстана в фотографиях». Алматы, 2015. 135 с.
27. Шарпило В.П. Паразитические черви пресмыкающихся фауны СССР. Киев: Наукова думка, 1976. 287 с.
28. Шилов И.А. Эколого-физиологические основы популяционных отношений у животных. М.: Изд-во МГУ, 1977. 261 с.
29. Щербак Н.Н. Разноцветная ящурка. Коллективная монография. Киев: «Наукова думка», 1993. 240 с.
30. Alcantara E.P., Ferreira-Silva C., Sousa J.G.G., Ávila R.W., Morais D.H. Ecology and parasitism of the lizard Tropidurus jaguaribanus (Squamata: Tropiduridae) from northeastern Brazil // Phyllomedusa. 2018. V. 17, No 2. P. 195–210.
31. Amorim D.M., Ávila R.W. Infection Patterns of Helminths in Norops Brasiliensis (Squamata, Dactyloidae) from a Humid Forest, Northeastern Brazil and their Relation with Body Mass, Sex, Host Size, and Season // Helminthologia. 2019. V. 56, No 2. P. 168–174. https://doi.org/10.2478/helm-2019-0011.
32. Barbosa D. Relationships between helminth communities and host traits in Podarcis bocagei and Podarcis carbonelli from NW Portugal // Italian Journal of Zoology. 2006. V. 73, No. 3. P. 213–217.
33. Bezerra C.H., Ávila R.W., Passos D.C., Zanchi-Silva D., Galdino C.A.B. Levels of helminth infections in the flat lizard Tropidurus semitaeniatus from north-eastern Brazil // J Helminthol. 2015. V. 90, No. 6. P. 779–783. https://doi.org/10.1017/S0022149X15000826.
34. Bezerra C.H., Pinheiro L.T., Melo de G.C., Zanchi-Silva D., Queiroz de M.S., Anjos dos L.A., Harris D.J., Borges-Nojosa D.M. Assessing the influence of geographic distance in parasite communities of an exotic lizard // Acta Parasitologica. 2016. V. 61, No. 1, P. 136–143. https://doi.org/10.1515/ap-2016-0018.
35. Bezerra C.H., Pinheiro L.T., Melo G.C., Braga R.R., Anjos L.A., Borges-Nojosa D.M. How epidemiological patterns shift across populations in an exotic lizard. // J Helminthol. 2019. V. 94, No. 97. P. 1–6. https://doi.org/10.1017/S0022149X19000907.
36. Birlik S., Yıldırımhan H.S., Kumlutas Y., Candan K., Ilgaz C. The first helminth study on Brandt's Persian lizard Iranolacerta brandtii (De Filippi, 1863) (Squamata: Lacertidae) from Van Province, Turkey // Helminthologia. 2017. V. 54, No. 2. P. 74–178.
37. Brown D.S., Symondson W.O. Sex and age-biased nematode prevalence in reptiles // Mol. Ecol. 2014. V. 23, No 15. P.3890–3899. https://doi.org/10.1111/mec.12688.
38. Er-Rguibi O., Laghzaoui E.M., Aglagane A., Kimdil L., Abbad A., El Mouden E.H. Determinants of prevalence and co-infestation by ecto- and endoparasites in the Atlas day gecko, Quedenfeldtia trachyblepharus, an endemic species of Morocco // Parasitol Res. 2021. V. 120, No. 7. P. 2543–2556. https://doi.org/10.1007/s00436-021-07120-z.
39. Fenner A.L., Bull C.M. The impact of nematode parasites on the behaviour of an Australian lizard, the gidgee skink Egernia stokesii // Ecological Research. 2008. V. 23. No. 5. P. 897– 903. https://doi.org/10.1007/s11284-007-0453-1.
40. Galdino C.A., Ávila R.W., Bezerra C.H., Passos D.C., Melo G.C., Zanchi-Silva D. Helminths infection patterns in a lizard (Tropidurus hispidus) population from a semiarid Neotropical area: associations between female reproductive allocation and parasite loads // J. Parasitol. 2014. V. 100. No. 6. P. 864–867.
41. Gyawali P.S., Khanal S., Shrestha B. Intestinal helminth fauna in sleepy lizard (Tiliqua rugosa) in Australia // Inter J Vet Sci. 2013. V. 2, No. 1. P. 17–20.
42. Le Galliard J-F., Fitze P.S., Ferrière R., Clobert J. Sex ratio bias, male aggression, and population collapse in lizards // PNAS. 2005. V. 102, No. 50. P. 18231–18236; https://doi.org/10.1073/pnas.0505172102.
43. Leung T.L.F., Koprivnikar J. Your infections are what you eat: How host ecology shapes the helminth parasite communities of lizards // J Anim Ecol. 2019. V. 88, No. 3. P. 416–426. https://doi.org/10.1111/1365-2656.12934.
44. Martin J.E., Roca V. Helminths of the Atlantic lizard, Gallotia atlantica (Reptilia: Lacertidae), in the Canary Islands (eastern Atlantic): composition and structure of component communities // Acta Parasitologica. 2005. V. 50, No. 1. P. 85–89.
45. Mayhew W.W. Biology of the granite spring lizard, Sceloporus orcutti // Amer. Midl. Nat. 1963. V. 69, No 2. P. 310–327.
46. Moritz C., McCallum H., Donnellan S., Roberts J.D. Parasite loads in parthenogenetic and sexual lizards (Heteronotia binoei). Support for the Red Queen hypothesis // Proceedings of the Royal Society B. 1991. V. 244, No. 1310. P. 145–149. https://doi.org/10.1098/rspb.1991.0063.
47. Rataj A.V., Lindtner-Knific R., Vlahović K., Mavri U., Dovč A. Parasites in pet reptiles // Acta Vet Scand. 2011. V. 53, No. 33. P. 1–20. https://doi.org/10.1186/1751-0147-53-33.
48. Roca V., Carretero M.A., Marques A., Barbosa D., Galdón M.A. Relationships between helminth communities and host traits in Podarcis bocagei and Podarcis carbonelli from NW Portugal // Italian Journal of Zoology. 2006. V. 73, No. 3. P. 213–217. https://doi.org/10.1080/11250000600727634.
49. Roca V., Jorge F., Ilgaz C., Kumlutas Y., Durmus S.H., Carretero M.A. Are the helminth communities from unisexual and bisexual lizards different? Evidence from gastrointestinal parasites of Darevskia spp. in Turkey // Acta Zoologica Academiae Scientiarum Hungaricae. 2015. V. 61, No. 3. P. 279–288.
50. Teixeira A.A. M., Silva R.J., Brito S.V., Teles D.A., Araujo-Filho J.A., Franzini L.D., Santana D.O., Almeida W.O., Mesquita D.O. Helminths Infecting Dryadosaura Nordestina (Squamata: Gymnophthalmidae) from Atlantic Forest, Northeastern Brazil // Helminthologia. 2018. V. 55, No. 4. P. 286–291. https://doi.org/10.2478/helm-2018-0026.
51. Tinkle D.W., Woodward D.W. Relative movements of lizards in natural populations as determined from receptive radii // Ecology. 1967. V. 48, No. 1. P. 166–168.