1.0 Введение: Система комплемента кишечника как новый рубеж в терапии
Понимание системы комплемента переживает фундаментальный сдвиг: от традиционного взгляда на нее как на системный, преимущественно печеночный механизм врожденного иммунитета к признанию ее роли как локально регулируемого медиатора в тканях. Этот парадигмальный сдвиг особенно важен для кишечника, где локальная система комплемента представляет собой важнейший, но до недавнего времени недостаточно изученный компонент иммунитета слизистых. Она является ключевым медиатором, регулирующим взаимодействие между хозяином и микробиотой и определяющим траекторию развития заболеваний. Данный документ предназначен для оценки этого нового рубежа с точки зрения разработки терапевтических препаратов для биотехнологической и фармацевтической отраслей, систематизируя последние научные достижения в этой области.
Система комплемента кишечника выполняет двойственную роль. С одной стороны, она поддерживает иммунный толеранс по отношению к комменсальным микроорганизмам и обеспечивает первую линию защиты от кишечных инфекций. С другой стороны, ее дисрегуляция становится мощным драйвером патологических процессов. Чрезмерная или неконтролируемая активация комплемента способствует хроническому воспалению и повреждению тканей, играя ключевую роль в патогенезе таких тяжелых состояний, как воспалительные заболевания кишечника (ВЗК) и колоректальный рак (КРР).
Цель — проанализировать сложные механизмы, с помощью которых система комплемента кишечника влияет на развитие и прогрессирование ВЗК и КРР, и на основе этого анализа определить наиболее перспективные терапевтические стратегии, основанные на модуляции путей комплемента.
Для достижения этой цели необходимо прежде всего понять фундаментальные отличия локальной системы комплемента кишечника от ее системного аналога. Далее мы подробно рассмотрим ее уникальные характеристики, которые и открывают возможности для разработки таргетных и безопасных терапевтических подходов.
2.0 Уникальная архитектура и функции системы комплемента в кишечнике
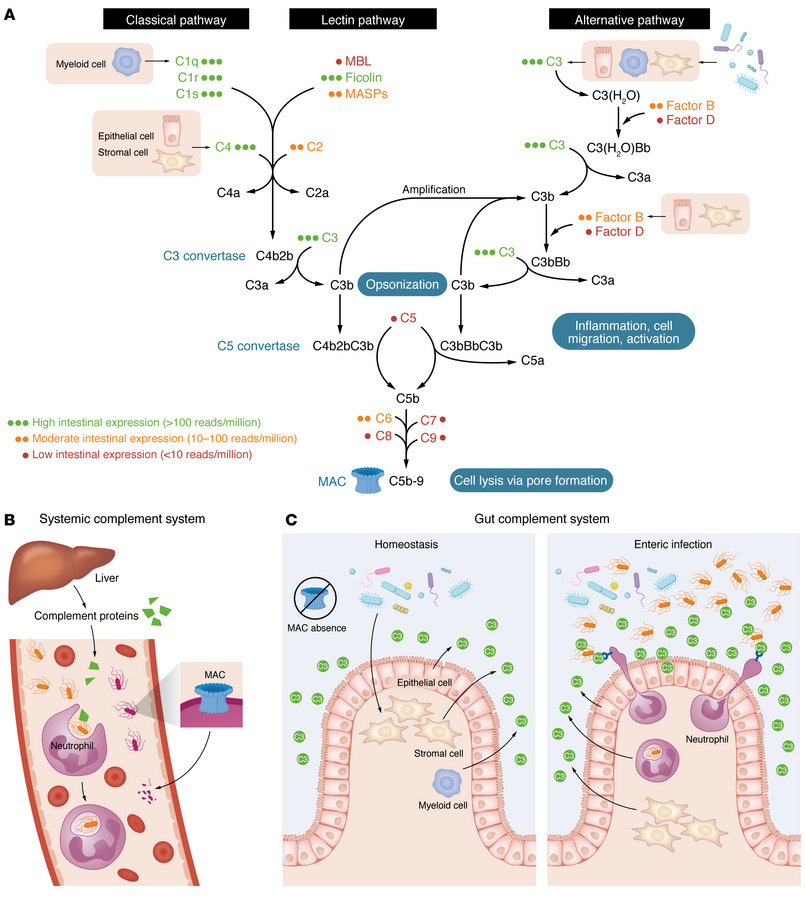
Стратегическое понимание уникальных адаптаций системы комплемента в кишечнике по сравнению с системным комплементом является основополагающим для разработки селективных терапевтических подходов. Эти отличия, сформированные в ходе эволюции для поддержания баланса в богатой микробной среде, определяют как ее защитные функции, так и ее роль в патологии. Ключевые особенности включают локальный синтез компонентов, отсутствие терминального каскада и доминирование альтернативного пути активации.
2.1 Локальный синтез компонентов комплемента
В отличие от системного комплемента, компоненты которого синтезируются преимущественно гепатоцитами, в кишечнике существует автономная система их производства. Эпителиальные клетки, стромальные клетки (в частности, фибробласты) и миелоидные популяции являются основными производителями центрального компонента C3, а макрофаги, локализованные в мышечной оболочке, — C1q. Компоненты классического (C4) и альтернативного (Фактор B) путей также синтезируются локально. Эта децентрализованная продукция обеспечивает быстрый и локализованный ответ на микробные сигналы.
Компонент / Основные клеточные источники / Ключевые функции в кишечнике
C1q / Макрофаги, эпителиальные клетки / Регуляция моторики кишечника через нейрональную экспрессию генов; ограниченная роль в иммунной защите
C3 / Эпителиальные клетки, стромальные клетки, миелоидные клетки / Опсонизация бактерий, стимуляция фагоцитоза, регуляция аутофагии
C4 / Эпителиальные клетки, стромальные клетки / Активация классического пути
Фактор B / Эпителиальные клетки, стромальные клетки / Активация альтернативного пути
Важно отметить, что уровни C3 в просвете кишечника напрямую регулируются микробиотой. Исследования на безмикробных мышах и животных, получавших антибиотики широкого спектра действия, демонстрируют значительное снижение концентрации C3 в фекалиях, что подтверждает ключевую роль микробных сигналов в поддержании его гомеостатического уровня.
2.2 Функциональные особенности: Отсутствие терминального каскада
Ключевым отличием и важнейшей адаптацией кишечной системы комплемента является отсутствие экспрессии терминальных компонентов (C5–C9). Эти белки необходимы для формирования мембраноатакующего комплекса (МАК) — поры, которая вызывает прямой лизис клеток-мишеней. Отсутствие МАК в кишечнике является критическим механизмом для поддержания иммунной толерантности к огромному сообществу комменсальных бактерий, предотвращая их массовое уничтожение.
Вследствие этого основной механизм защиты в кишечнике — это не прямой лизис, а опсонизация (покрытие бактерий фрагментами комплемента, такими как C3b) с последующим фагоцитозом рекрутированными иммунными клетками. Основным путем активации комплемента в кишечнике является альтернативный путь, что обеспечивает мощный механизм амплификации сигнала. Примечательно, что такая архитектура напоминает защитные системы древних книдарий и представляет собой древний, нелитический защитный механизм, идеально приспособленный для слизистых оболочек.
Уникальная архитектура системы комплемента кишечника, ориентированная на опсонизацию, а не на лизис, требует наличия специфических клеточных рецепторов для распознавания фрагментов комплемента и регуляторных белков для предотвращения повреждения собственных тканей.
3.0 Клеточные механизмы: Рецепторы и регуляторы комплемента в кишечнике
Эффективность и контроль системы комплемента полностью зависят от сложной сети клеточных рецепторов, которые инициируют сигналы, и мембраносвязанных регуляторных белков, которые защищают клетки хозяина от повреждения. Понимание паттернов их экспрессии в различных клеточных популяциях кишечника имеет решающее значение для идентификации высокоспецифичных терапевтических мишеней.
3.1 Экспрессия и функции рецепторов комплемента
Рецепторы комплемента делятся на две основные группы: рецепторы анафилатоксинов (C3aR, C5aR1) и опсониновые рецепторы (CR1, CR3, CRIg и др.).
• Рецепторы анафилатоксинов: В собственной пластинке слизистой оболочки кишечника рецептор C3aR экспрессируется на макрофагах, дендритных клетках (ДК), врожденных лимфоидных клетках 3 типа (ILC3), а также на Lgr5+ стволовых клетках кишечника, где он стимулирует их экспансию. Примечательно, что в макрофагах собственной пластинки C3aR преимущественно локализован внутриклеточно, что указывает на участие внутриклеточного комплемента («комплесомы») в регуляции воспаления. Рецептор C5aR1 имеет более ограниченную экспрессию, преимущественно на макрофагах и лизоцим-экспрессирующих ДК, где он усиливает перекрестную презентацию антигенов и иммунитет CD8+ Т-клеток, хотя его экспрессия также сообщалась на эпителиальных клеточных линиях (хотя эти данные получены на трансформированных клеточных линиях и не подтверждены на первичных эпителиальных клетках).
• Опсониновые рецепторы: Эти рецепторы широко представлены на иммунных клетках кишечника. Особый интерес представляет CRIg (VSIG4), который экспрессируется на тканерезидентных макрофагах. Его экспрессия постнатально регулируется сигналами от микробиоты и значительно снижается при ее истощении антибиотиками. Эти CRIg+ макрофаги играют важную роль в поддержании гомеостаза, в частности, путем клиренса внеклеточных везикул микробного происхождения.
3.2 Роль комплемент-регуляторных белков (КРБ)
Для предотвращения атаки на собственные клетки на поверхности эпителия экспрессируются комплемент-регуляторные белки (КРБ), такие как CD55, CD59 и CD46. Их экспрессия динамически изменяется при заболеваниях.
Белок / Локализация в кишечнике / Патологическая регуляция (ВЗК/КРР)
CD55 (DAF) / Апикальная поверхность эпителия крипт тонкого кишечника / Значительно повышена при ВЗК (БК, ЯК) и КРР
CD59 (Protectin) / Апикальная поверхность эпителия толстого кишечника / Не изменена или снижена в толстом кишечнике при ЯК и БК; значительно повышена при КРР
CD46 (MCP) / Базолатеральная поверхность эпителия по всему кишечнику / Повышена при КРР
Анализ показывает, что при воспалении и раке экспрессия КРБ претерпевает значительные изменения. Например, при КРР опухолевые клетки активно повышают экспрессию CD55 и CD59, что является ключевым механизмом уклонения от комплемент-опосредованного лизиса и коррелирует с плохим прогнозом для пациентов.
Дисбаланс между активацией комплемента и его регуляцией является центральным элементом патогенеза хронических воспалительных заболеваний, что наиболее ярко проявляется при ВЗК.
4.0 Дисфункция комплемента при воспалительных заболеваниях кишечника (ВЗК)
При ВЗК система комплемента демонстрирует свою двойственную природу: с одной стороны, она необходима для защиты от патогенов и восстановления тканей, но с другой — ее чрезмерная и хроническая активация усугубляет воспаление и приводит к значительному повреждению слизистой оболочки.
Накоплены убедительные доказательства как системной, так и локальной дисрегуляции комплемента у пациентов с ВЗК. Ключевые факты включают:
• Повышенный синтез C3: У пациентов с ВЗК наблюдаются значительно более высокие показатели синтеза C3 по сравнению со здоровыми людьми.
• Различия между БК и ЯК: Уровни C3 и C1q в сыворотке крови выше у пациентов с болезнью Крона (БК), чем у пациентов с язвенным колитом (ЯК).
• Локальное производство: В еюнальных секретах пациентов с БК обнаружены повышенные уровни C3 и C4, что указывает на их локальный синтез в тонком кишечнике.
Пути активации комплемента также различаются при двух основных формах ВЗК. При язвенном колите преобладает классический путь, что подтверждается отложением IgG1 и C3b на эпителии. В то же время при болезни Крона доминирует альтернативный путь, о чем свидетельствует отложение C3b в отсутствие IgG или C1q.
Эта аберрантная активация комплемента тесно связана с дисбиозом кишечной микробиоты. Снижение микробного разнообразия и рост патобионтов, характерные для ВЗК, выступают вероятным механистическим триггером, модулирующим локальные уровни и статус активации C3. Это запускает порочный круг, в котором измененная микробиота усиливает комплемент-опосредованное воспаление, а воспаление, в свою очередь, усугубляет дисбиоз.
Хроническое воспаление, поддерживаемое комплементом, создает провоспалительное микроокружение, способствующее злокачественной трансформации клеток, что напрямую подводит к анализу роли комплемента в развитии колоректального рака.
5.0 Двойственная роль комплемента в патогенезе колоректального рака (КРР)
В контексте колоректального рака система комплемента представляет собой центральный терапевтический парадокс. Ее эффекторные механизмы обладают как про-, так и противоопухолевым потенциалом: сигнальные пути анафилатоксинов (в частности, C5a/C5aR1) являются мощными драйверами онкогенеза, в то время как терминальный литический каскад (МАК) способен уничтожать раковые клетки. Это усложняет разработку терапевтических подходов и требует точного таргетирования конкретных путей.
5.1 Про-туморогенные эффекты сигнальных путей комплемента
Несмотря на способность комплемента лизировать раковые клетки, определенные его компоненты активно способствуют росту и распространению опухоли.
• Ось C5a/C5aR1 как драйвер онкогенеза: Данный сигнальный путь последовательно демонстрирует мощные про-опухолевые эффекты. Его активация:
◦ Усиливает пролиферацию, миграцию и инвазию раковых клеток.
◦ Привлекает в микроокружение опухоли миелоидные супрессорные клетки, которые подавляют противоопухолевую активность CD8+ Т-клеток.
◦ Стабилизирует онкогенный белок β-катенин в эпителиальных клетках.
◦ Способствует метастазированию опухоли в печень.
• Неоднозначная роль оси C3a/C3aR: Роль этого пути менее определена. С одной стороны, повышенные уровни C3a в сыворотке могут служить биомаркером КРР. С другой стороны, потеря функции C3aR в некоторых моделях способствует развитию опухоли за счет усиления инфильтрации провоспалительных иммунных клеток.
5.2 Механизмы уклонения опухоли от комплемент-опосредованной атаки
Чтобы выжить, клетки КРР выработали эффективные стратегии уклонения от уничтожения с помощью МАК. Основной механизм заключается в гиперэкспрессии на своей поверхности комплемент-регуляторных белков (КРБ).
Опухолевые клетки значительно повышают экспрессию CD55, CD46 и CD59. Эти белки ингибируют каскад комплемента на разных стадиях, предотвращая формирование летального МАК на мембране раковой клетки.
Клиническая значимость этого механизма огромна: высокая экспрессия КРБ напрямую коррелирует с плохим прогнозом у пациентов с КРР. Это делает данные белки не только прогностическими маркерами, но и привлекательными терапевтическими мишенями, блокирование которых может восстановить способность иммунной системы атаковать опухоль.
Глубокое понимание про-туморогенных путей и механизмов иммунного уклонения открывает конкретные и целенаправленные возможности для терапевтического вмешательства, которые необходимо систематизировать для дальнейшей разработки.
6.0 Определение терапевтических стратегий: Нацеливание на пути комплемента
Данный раздел синтезирует представленный выше анализ для определения конкретных и многообещающих терапевтических стратегий, представляющих интерес для фармацевтической разработки. Эти подходы направлены на восстановление гомеостаза и преодоление патологических механизмов, опосредованных комплементом при ВЗК и КРР.
1. Ингибирование оси C5a/C5aR1 при КРР
◦ Заболевание-мишень: КРР.
◦ Обоснование: Учитывая последовательную про-туморогенную роль оси C5a/C5aR1, ее блокирование является одной из наиболее перспективных стратегий. Это может одновременно подавить пролиферацию и метастазирование опухолевых клеток, а также преодолеть иммуносупрессивное микроокружение опухоли. Ключевое преимущество этого подхода заключается в его многоцелевом действии как на опухолевые клетки, так и на иммунный микроокружение.
2. Модуляция C3 при ВЗК
◦ Заболевание-мишень: ВЗК.
◦ Обоснование: Поскольку чрезмерная активация C3 является центральным звеном хронического воспаления при ВЗК, его таргетирование обладает высоким терапевтическим потенциалом. Основная задача для разработчиков — найти «узкое терапевтическое окно», позволяющее снизить патологическое воспаление, не компрометируя при этом ключевую роль C3 в защите от кишечных патогенов.
3. Нацеливание на комплемент-регуляторные белки (КРБ) при КРР
◦ Заболевание-мишень: КРР.
◦ Обоснование: Использование моноклональных антител против КРБ (CD55, CD46, CD59) может демаскировать опухоль, восстанавливая комплемент-опосредованный лизис. Этот подход имеет значительный потенциал для использования в комбинированных терапиях, например, с ингибиторами контрольных точек, для сенсибилизации опухолей к иммунной атаке.
4. Микробиота-опосредованная модуляция комплемента
◦ Заболевание-мишень: ВЗК.
◦ Обоснование: Эта стратегия представляет собой косвенный, но физиологически обоснованный подход. Вместо прямого фармакологического подавления, модуляция микробиома (например, с помощью пробиотиков нового поколения) нацелена на восстановление гомеостатической регуляции C3. Это можно рассматривать как «хозяин-центрированный подход к микробной проблеме», обладающий потенциалом для долгосрочной ремиссии.
Реализация этих многообещающих стратегий потребует дальнейших глубоких исследований для точного определения пространственных, клеточных и молекулярных детерминант, управляющих активностью комплемента в сложной экосистеме кишечника.
7.0 Заключение и будущие перспективы
Система комплемента кишечника, долгое время остававшаяся в тени своего системного аналога, сегодня предстает как критический медиатор здоровья и заболеваний желудочно-кишечного тракта. Ее уникальная архитектура, характеризующаяся локальным синтезом, отсутствием терминального литического каскада и тесной связью с микробиотой, открывает новые терапевтические возможности для лечения ВЗК и КРР. Дисрегуляция этого тонко настроенного механизма превращает его из защитника в мощный драйвер хронического воспаления и онкогенеза.
Будущие направления исследований должны быть сосредоточены на расшифровке точных сигналов — как микробного, так и хозяйского происхождения — которые управляют продукцией и активацией комплемента в различных компартментах кишечника. Углубленное понимание этих детерминант будет иметь решающее значение для успешного преобразования фундаментальных знаний в эффективные и безопасные клинические вмешательства.
Точное нацеливание на конкретные пути комплемента обещает появление нового класса терапевтических средств, знаменующих переход от широкой иммуносупрессии к высокоточной модуляции, направленной на восстановление гомеостаза слизистых оболочек. Такие подходы способны кардинально улучшить исходы при широком спектре желудочно-кишечных заболеваний.