Вступление
Хотя мезозой и называют эпохой динозавров, таковой он стал далеко не сразу, ведь предкам динозавров пришлось пройти долгий путь от мелких животных до величественных гигантов. Динозавры, как и птерозавры, принадлежат к группе животных под названием Авеметатарзалии («птичьи предплюсны») которые впервые появляются в среднем триасе. О них собственно и пойдёт речь далее.
Авеметатарзалии (особенности)
Авеметатарзалии являются второй ветвью архозавров (первая это псевдозухии) которая содержит внутри себя орнитодир (предков и ближайших родственников, а также самих птерозавров и динозавров), а также афанозаврий («скрытых ящериц») и неклассифицированных таксонов (таких как мамбачитон).

Основополагающей характеристикой авеметатарзалий являются «усовершенствованные мезатарзальные» лодыжки, для которых характерны большой астрагал и маленькая пяточная кость. Такая ориентация лодыжки работает на одном шарнире, обеспечивая лучшую подвижность. Вероятно, в результате этого изменения общий предок птицетазовых имел вертикальную, двуногую осанку, при которой ноги располагались вертикально, как у млекопитающих. Авеметатарзалии, как правило, были более лёгкого телосложения, чем крокодиловые архозавры. У них были более мелкие головы и, как правило, полное отсутствие остеодерм.
Самой примитивной и базальной группой авеметатарзалий были афанозаврии, о которых пойдёт речь ниже.
Афанозаврии
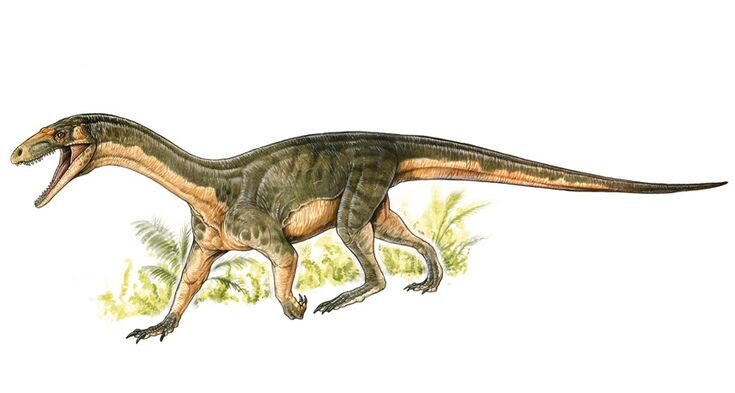
Афанозаврии («скрытые ящерицы») – базальные члены клады авеметатарзалий, процветавшие в среднем триасе. Представители этой группы были лёгкими и среднего размера рептилиями. У них не было никаких приспособлений для передвижения на двух ногах, что стало гораздо более распространённым у ящеротазовых динозавров. Кроме того, пропорции их ног указывают на то, что они не были способны к длительному бегу, а значит, были медленными по меркам ящеротазовых динозавров.
Череп
Для группы в целом известно очень мало черепных материалов. Единственные кости черепа, которые можно с уверенностью отнести к этой группе, — это несколько фрагментов крыловидной и заглазничной костей, принадлежащих ярасухусу, а также несколько фрагментов, которые, как считается, принадлежат телеократеру. К этим костям относятся верхняя челюсть (зубчатая кость в средней части морды), лобная кость (часть свода черепа над глазами) и квадратная кость (часть челюстного сустава черепа). Хотя эти фрагменты затрудняют реконструкцию черепа афанозавров, они демонстрируют несколько примечательных особенностей. Например, форма верхней челюсти показывает, что у афанозавров была предглазничная фенестра — большое отверстие на морде прямо перед глазами. В сочетании с предглазничной впадиной (разрушенной областью кости, которая окружала глазницу) это указывает на то, что афанозавры принадлежали к группе архозавров. На нижнем крае верхней челюсти также сохранился частично прорезавшийся зуб. Этот зуб был уплощён с боков, слегка изогнут назад и зазубрен по переднему краю. Эти особенности зубов указывают на то, что афанозавры были плотоядными, как и многие мясоядные рептилии (включая динозавры-тероподы, такие как велоцираптор и дейноних) имели те же особенности. На переднем крае верхней челюсти также есть небольшая ямка, похожая на ямку у некоторых силезаврид. В задней части лобной части имелась круглая неглубокая ямка, известная как надвисочная ямка. Раньше считалось, что только у динозавров были надглазничные углубления, но их наличие у афанозавров (и асилизавра, силезаврида) показывает, что они были характерны для многих авеметатарзалий. В целом, известные черепа афанозавров не имели уникальных особенностей, а это значит, что для характеристики группы пришлось бы использовать остальную часть скелета.
Позвонки
Афанозавры имеют множество отличительных особенностей в строении шейных позвонков (шейных позвоночников). Шейные позвонки очень длинные по сравнению с другими ранними авеметатарзалиями. Как и у большинства других рептилий, позвонки состоят из основного тела (центральной части) и похожего на пластину неврального отростка, выступающего сверху. В передних шейных позвонках (позвонках в передней части шеи) по нижней стороне центральной части проходит пара низких гребней. Эти гребни разделены широкой областью с другими, более мелкими гребнями, что делает центр тела примерно прямоугольным в поперечном сечении. Нервные отростки шейных позвонков также уникальны у афанозавров. Они имеют топористую форму, с передними краями, которые сужаются к острию и резко нависают над центром, по крайней мере, в передней и средней частях шеи. Верхний край нервного отростка тонкий и похожий на лезвие, но область непосредственно под краем приобретает грубую текстуру и образует низкий округлый гребень. Все эти особенности уникальны для афанозавров.
Как и у других рептилий, у афанозавров позвонки также имеют небольшие структуры, которые соединяются либо с другими позвонками, либо с рёбрами, которые прикрепляются к каждому позвонку. Структуры, которые соединяются с позвонками спереди, называются презигапофизами, а те, которые соединяются с позвонками сзади, называются постзигапофизами. Структуры, которые соединяются с рёбрами, также имеют разные названия. У большинства архозавров головки рёбер двузубчатые. В результате на боковой стороне каждого позвонка есть две области для соединения с ребром: диафиз в верхней части тела позвонка и парафиз в нижней части. Однако некоторые шейные рёбра у афанозавров очень необычны, так как имеют трёхконечную головку, хотя такая особенность встречается только у рёбер в основании шеи. В сочетании с этой особенностью позвонки в этой области имеют выступ для третьего отростка прямо над парапофизом, который иногда называют «разделенным парапофизом». Единственными другими архозаврами с такой особенностью были попосавроиды, что объясняет, почему ярасуха в прошлом ошибочно принимали за попосавроида.
Помимо этих уникальных для авеметатарзалий особенностей, у афанозавров есть ещё несколько черт, присущих другим группам. В позвонках передней и средней части шеи постзигапофизы имеют дополнительные небольшие отростки прямо над сочленяющимися пластинами. Эти дополнительные отростки называются эпипофизами и распространены у динозавров, но, вероятно, развились независимо, поскольку отсутствуют у других групп авеметатарзалий. Позвонки тела имеют другой тип вторичной структуры. Небольшая структура (гипофен) под постзигапофизами вставляется в губу (гипантрум) между презигапофизами следующего позвонка, образуя дополнительные сочленения, которые помогают зигапофизам. Эти сочленения гипофен-гипантрум присутствуют у зауроподовых динозавров, а также у раизахид, и часто считается, что они помогают сделать позвоночник более жёстким.
Передние конечности
У афанозавров есть несколько характерных черт плечевой кости (предплечья). Эта кость была прочной, тонкой, если смотреть сбоку, но широкой, если смотреть спереди. При переднем (фронтальном) взгляде ее середина была ущемлена, в то время как проксимальный (ближний) и дистальный (дальний) концы были широкими, что придавало кости форму песочных часов. Край верхней части плечевой кости, обращенный в сторону от тела, имеет закругленный гребень, известный как дельтопекторальный гребень. Этот гребень направлен вперёд и довольно вытянут, занимая примерно треть длины кости. В целом плечевая кость афанозавров очень похожа на плечевую кость завропод и ньясазавра, неопределённого раннего динозавра или родственника динозавров. Рука в целом была крепкой и немного короче ноги, но только плечевая кость обладала уникальными особенностями. Рука представителей этой группы в основном неизвестна, но предположительно она была маленькой и пятипалой, как у большинства архозавров (за исключением специализированных форм, таких как птерозавры или динозавры-тероподы).
Бедро и задние конечности
Тазовый пояс
Таз (бедро) афанозавров имеет много общего с тазом ранних динозавров и силезаврид, а также с тазом неродственных им попозаврид. Большинство этих черт можно обнаружить в седалищной кости, которая представляет собой плоскую кость, образующую нижнюю заднюю часть бедра. Например, каждая седалищная кость (с обеих сторон бедра) соприкасается друг с другом по средней линии бедра. Этот контакт очень обширный, хотя кости и не срастаются полностью из-за того, что контакт не доходит до верхнего края каждой кости. В отличие от них, у птерозавров, лагерпетид и маразухов (других птицетазовых) седалищные кости лишь слегка соприкасаются в средней части каждой кости. Кончик седалищной кости также округлый и полутреугольный в поперечном сечении, при этом латеральная (внешняя) сторона каждой седалищной кости сужается к нижнему краю кости, а медиальная (внутренняя) сторона плоская и соприкасается с другой седалищной костью. У потозавроидов и динозавров седалищные кости также округлые, но не имеют полутреугольной формы, которая также известна у асилизавра. На верхней части седалищной кости есть бороздка. В отличие от динозавров, у афанозавров есть вертлужная впадина (тазобедренный сустав), которая закрыта костью, хотя, возможно, небольшая часть была открыта в соответствии с выемкой в том месте, где седалищная кость соприкасается с подвздошной костью (верхней частью бедра).
Нога
Тонкая бедренная кость (тазовая кость) афанозавров обладает характерным набором признаков, по которым можно определить принадлежность к группе. Проксимальная (ближняя) поверхность кости, которая соединяется с вертлужной впадиной, имеет глубокую борозду, а не просто плоскую суставную поверхность. Кроме того, дистальная (дальняя) суставная поверхность кости, которая соединяется с костями голени, вогнутая. Проксимальная часть бедренной кости также имеет несколько бугорков (выступов) на внешнем или внутреннем крае кости. У многих птицетазовых динозавров на внутреннем крае есть два таких выступа: небольшой переднемедиальный выступ спереди и более крупный заднемедиальный выступ сзади. Однако у афанозавров, по-видимому, полностью отсутствует (или никогда не было) переднемедиальный выступ. Это почти беспрецедентно для архозавров, но похоже на то, что происходит у родственников архозавров таких как эупаркерия.
На внутренней части кости, примерно на четверть длины стержня, имеется небольшой выступ. Этот гребень, называемый четвертым вертелом, является местом прикрепления M. caudofemoralis, мышцы хвоста, которая помогает отводить задние конечности. Шрам на переднебоковом (переднем и наружном) крае бедренной кости, возможно, прикрепился к M. iliotrochantericus caudalis, мышце, которая соединяется с бедром и помогает стабилизировать бедро. Этот конкретный шрам может быть тем же самым, что и передний (или меньший) вертел, специфическая структура, присутствующая у динозавров и их близких родственников. Другой шрам расположен несколько дальше назад на кости и ниже на стержне. Этот шрам, возможно, прикрепился к M. iliofemoralis externus, мышце, которая играет ту же роль, что и M. iliotrochantericus caudalis. Точно так же его предполагаемым эквивалентом у динозавров является структура, известная как вертельный выступ. Афанозавры уникальны среди других птицетазовых тем, что эти две шпоры отделены друг от друга. У более развитых птицетазовых, таких как динозавры, эти две структуры и соответствующие им мышцы сливаются, и это состояние сохраняется у современных птиц.
Тонкие большеберцовая и малоберцовая кости (кости голени) афанозавров не обладают уникальными чертами в той же степени, что бедренная кость. Однако они также короче бедренной кости. Такие пропорции редки среди ранних авеметатарсалиев, но более распространены среди псевдозухов и неархозавровых архозавриформ. Короткая голень обратно пропорциональна способностям к бегу, указывая на то, что афанозавры были не такими быстрыми и проворными, как более продвинутые представители авеметатарзалий.
Лодыжка
У двух разных афанозавров (ярасухуса и телеократера) сохранилась пяточная кость, также известная как таранная кость. У большинства авеметатарзалий пяточная кость простая и прочно соединена с большой костью, известной как астрагал, расположенной рядом с ней. Такой тип пяточной кости, известный как «продвинутая мезотарзальная» форма, обеспечивает большую устойчивость, но меньшую гибкость стопы, поскольку разные кости голеностопного сустава не могут сгибаться относительно друг друга. Псевдозухии (в том числе современные крокодилы), а также крокодилоподобные фитозавры имеют другую конфигурацию: пяточная кость у них намного больше и сложнее, она соединяется с астрагалом с помощью сустава, который позволяет им двигаться. Такая конфигурация называется «крокодильей» лодыжкой, а рептилии, у которых она есть, называются круротаранными. Некоторые недавние исследования показали, что фитозавры на самом деле не являются архозаврами, а являются их близкими родственниками.Это указывает на то, что «крокодильи» лодыжки были плезиоморфным (базовым) состоянием у первых архозавров, а «усовершенствованные мезотарсальные» лодыжки появились только позже в пределах авеметатарзалий, а не у основания группы.
Пяточная кость афанозавров подтверждает эту идею, поскольку она больше похожа на «нормальную» для крокодилов лодыжку, чем на «продвинутую» мезотарсальную лодыжку. Пяточная кость расположена с внешней стороны лодыжки, её передний или внутренний край соединяется с астрагалом, верхняя поверхность соединяется с малоберцовой костью, а нижняя — с четвёртой плюсневой костью (малой костью стопы). У афанозавров суставная впадина для астрагала вогнутая, а соединение с малоберцовой костью имеет форму округлого купола. Это характерные черты «крокодильей» лодыжки. Кроме того, задняя часть пяточной кости имеет цилиндрическую структуру, известную как пяточный бугор. Хотя у афанозавров этот бугор меньше, чем у псевдозухий, он всё равно намного больше, чем у других птицетазовых, у большинства из которых он вообще отсутствует. У некоторых динозаврообразных также есть небольшие костные наросты на пяточной кости, хотя у афанозавров наросты крупнее и круглее, чем у этих таксонов (маразух и несколько базальных силезаврид). В поперечном сечении пяточные кости афанозавров имеют овальную форму, они выше, чем шире. В этой группе сохранилось мало фрагментов ступней, известно лишь несколько фаланг (костей пальцев) и плюсневых костей (основных удлинённых костей ступни). Судя по длине сохранившихся плюсневых костей, ступня, вероятно, была довольно длинной.
Афанозавры распространились по всей Пангее заняв место практически во всех тогдашних экосистемах, так например известен донгузух из Донгузской формации в России, телеократер из формации Манда в Танзании, яразух из формации Йерапалли в Индии и спондилосома из формации Санта-Мария в Бразилии.