Оплодотворение представляет собой процесс объединения ядра сперматозоида отца с ядром яйцеклетки матери, что приводит к образованию первичного ядра зародыша. Суть этого явления заключается в слиянии генетического материала двух различных половых клеток — гамет, каждая из которых содержит половину набора хромосом, характерного для конкретного вида. У микроорганизмов и простейших наиболее простая форма оплодотворения заключается в обмене генетической информацией между двумя клетками.
Первым значимым этапом в процессе оплодотворения является слияние мембран двух гамет, что создает канал для переноса генетического материала из одной клетки в другую. В случае с развитыми растениями оплодотворению предшествует опыление, при котором пыльца попадает на женскую гамету или макроспору и вступает в контакт с ней. У развитых животных оплодотворение обычно сопровождается проникновением в яйцеклетку одного сперматозоида, после чего формируется клетка (зигота), способная к делению для развития новой особи.
Слияние двух гамет запускает ряд реакций в яйцеклетке, одна из которых изменяет мембрану, предотвращая проникновение большего количества сперматозоидов. Даже у видов с полиспермией, только одно сперматозоидное ядро фактически соединяется с ядром яйцеклетки. Основной результат оплодотворения — это активация яйцеклетки, которая инициирует её клеточное деление. Активация может происходить и без участия сперматозоида, например, при партеногенезе, под воздействием физических или химических факторов. Многие беспозвоночные, такие как тли, пчелы и коловратки, обычно размножаются партеногенезом.
У растений определенные химические вещества, выделяемые яйцеклеткой, могут привлекать сперматозоиды. У животных, за исключением, вероятно, некоторых книдарий, контакт между яйцеклетками и сперматозоидами чаще всего случаен. Желатиновые оболочки, окружающие яйца многих животных, могут задерживать сперматозоиды, что повышает вероятность успешного взаимодействия.
Яйца морских беспозвоночных, особенно иглокожих, служат классическими объектами для исследования оплодотворения. Их прозрачность и возможность фиксировать точное время оплодотворения делают их ценными для изучения живых клеток и проведения биохимических и молекулярных исследований. Яйца некоторых костистых рыб и амфибий также использовались для этих целей с положительными результатами.
Созревание яйца Созревание, или оогенез, представляет собой заключительный этап в формировании функциональных яйцеклеток, способных объединяться со сперматозоидом и вырабатывать реакции, препятствующие проникновению дополнительных сперматозоидов. Цитоплазма зрелой яйцеклетки также поддерживает процессы, приводящие к слиянию ядер сперматозоида и яйцеклетки и началу эмбрионального развития.
Поверхность яйца морского ежа
Некоторые элементы поверхности яйца морского ежа, особенно кортикальные гранулы, указывают на его зрелость. Эти гранулы, находящиеся под плазматической мембраной созревших яиц, имеют диаметр 0,8-1,0 микрон (0,0008-0,001 миллиметра) и окружены мембраной, схожей по структуре с плазматической мембраной, окружающей яйцо. Кортикальные гранулы формируются в аппарате Гольджи и перемещаются на поверхность созревающей яйцеклетки.
Поверхность яйца морского ежа обладает уникальной способностью по-разному преломлять свет в различных направлениях. Это явление, известное как двойное лучепреломление, указывает на определенную организацию молекул в поверхностных слоях. Поскольку такое преломление света возникает по мере созревания яйца, можно предположить, что особенности мембраны зрелого яйца связаны с упорядоченным расположением молекул. Зрелая яйцеклетка способна поддерживать формирование зиготного ядра, образующегося при слиянии ядер сперматозоида и яйцеклетки. В большинстве яйцеклеток процесс сокращения числа хромосом (мейоз) завершается только после оплодотворения. В таких случаях оплодотворяющий сперматозоид остается под поверхностью яйцеклетки до завершения мейоза, а потом происходят изменения и движения, приводящие к образованию зиготы.
Оболочки яиц
Поверхность большинства животных яйцеклеток окружена оболочками, которые могут быть мягкими и желатиновыми (у иглокожих и земноводных) либо плотными и толстыми (у рыб, насекомых и млекопитающих). Для того чтобы достичь поверхности яйцеклетки, сперматозоиды должны пробить эти оболочки, используя ферменты (биологические катализаторы), которые разрушают эти оболочки. В некоторых случаях (например, у рыб и насекомых) в оболочке имеется микропила, через который сперматозоид проникает в яйцеклетку.
Желеобразные оболочки яиц иглокожих и амфибий состоят из сложных углеводов, называемых сульфатированными мукополисахаридами. Оболочка яйцеклетки млекопитающих более сложная. Яйцо окружено толстой оболочкой, состоящей из углеводно-белкового комплекса под названием zona pellucida. Внешняя оболочка, corona radiata, состоит из множества слоев клеток и образована фолликулярными клетками, которые прилегают к яйцеклетке до ее выхода из фолликула яичника.
Когда-то считалось, что желеобразная оболочка яйцеклетки иглокожих содержит вещество (фертилизин), которое играет важную роль в установлении взаимодействия между сперматозоидом и яйцеклеткой, а также в активации последней. Однако ныне доказано, что фертилизин идентичен материалу желеобразной оболочки, а не веществу, постоянно выделяемому из нее. Тем не менее, существуют данные, подтверждающие, что оболочки яйцеклетки действительно участвуют в процессе оплодотворения: например, контакт сперматозоида с оболочкой вызывает акросомную реакцию.
Объединение сперматозоида и яйцеклетки
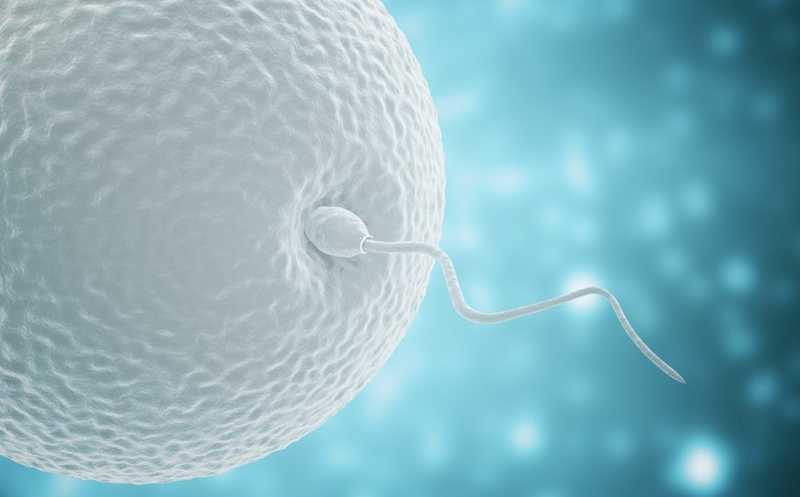
После контакта сперматозоида с внешними слоями яйцеклетки акросома — выступ на переднем конце сперматозоида — претерпевает определенные структурные изменения, что открывает путь для проникновения ядра сперматозоида. Акросомная реакция необходима для соединения сперматозоида с яйцеклеткой посредством слияния их плазматических мембран.
После соприкосновения сперматозоида с яйцеклеткой акросома проходит серию четко определенных структурных изменений. Внутренняя структура акросомы, акросомальный пузырек, разрывается, и плазматическая мембрана сперматозоида сливается с мембраной акросомального пузырька, образуя отверстие. В процессе этого гранула внутри акросомального пузырька растворяется, высвобождая вещество, называемое лизином, которое разрушает стекловидную оболочку яйцеклетки, позволяя сперматозоиду проникнуть внутрь.
Область акросомальной мембраны у отверстия соединяется с ядерной оболочкой сперматозоида, образуя выступ, который быстро удлиняется в тонкую трубку — акросомальный каналец. Этот каналец достигает поверхности яйцеклетки, устанавливая связь с ее плазматической мембраной. Образовавшийся каналец обеспечивает непрерывность между клетками и позволяет ядру сперматозоида пройти внутрь яйцеклетки. Другие структуры сперматозоида могут проникнуть внутрь яйцеклетки, включая среднюю часть и хвост, однако плазматическая и акросомальная мембраны сперматозоида внутрь не попадают.
Целые сперматозоиды, введенные в неоплодотворенные яйцеклетки, не способны вызвать активационную реакцию или слиться с ядром яйцеклетки. Во время проникновения ядра сперматозоида в яйцеклетку, его плазматическая мембрана разрушается; а в конце процесса целостность плазматической мембраны яйцеклетки восстанавливается. Это описание объединения сперматозоида с яйцеклеткой, впервые наблюдаемое у желудевого червя Saccoglossus (филум Enteropneusta), общей чертой характерно для большинства изученных до сих пор яиц.
Проходя через женские репродуктивные пути млекопитающих, сперматозоиды подвергаются важным физиологическим изменениям, известным как капацитация, что является ключевым моментом для их способности участвовать в оплодотворении. Эти изменения позволяют сперматозоидам проходить акросомную реакцию, проникать через оболочки яйцеклетки и достигать её внутренней части. Расщепление клеток внешней оболочки яйцеклетки (corona radiata) происходит под действием фермента гиалуронидазы, который разрушает гиалуроновую кислоту, соединяющую эти клетки. Гиалуронидаза может присутствовать в акросоме и высвобождается в ходе акросомной реакции, когда сперматозоид проходит через corona radiata. К моменту достижения сперматозоидом толстой оболочки яйцеклетки (zona pellucida), реакция уже активно развивается. Путь сперматозоида через zona pellucida формируется благодаря колебательным движениям его головки вблизи места прикрепления к оболочке.
При соприкосновении сперматозоида млекопитающего с поверхностью яйцеклетки, контакт происходит вдоль боковой части сперматозоида, что отличает его от других животных, у которых контакт осуществляется через кончик. Сперматозоид располагается параллельно поверхности яйцеклетки, и между их плазматическими мембранами образуется несколько точек слияния, что приводит к разрушению мембран и образованию множественных мелких везикул.
Специфический характер взаимодействия сперматозоида и яйцеклетки видоспецифичен, и оболочка яйцеклетки, зона пеллюцида, играет ключевую роль в связывании гамет. Обычно биохимический состав zona pellucida различается у различных видов, что позволяет ей взаимодействовать только со сперматозоидами своего же вида. Например, у иглокожих растворы желеобразной оболочки связывают исключительно сперматозоиды своего вида. Однако у иглокожих и амфибий небольшие повреждения поверхности яйца могут позволить оплодотворение сперматозоидами других видов (гетерологичное оплодотворение), что используется для получения гибридных личинок. Более того, удаление zona pellucida яйцеклетки может способствовать связыванию сперматозоидов и яйцеклеток разных видов.
Яйца асцидий, также именуемых морскими котиками и принадлежащих к подсемейству хордовых Tunicata, заключены в плотную оболочку под названием хорион. Между хорионом и яйцеклеткой располагаются клетки, называемые тестикулярными. У асцидий, которые одновременно содержат как мужские, так и женские половые органы, гаметы созревают синхронно, но самоплодотворение не происходит. Если же удалить хорион и тестикулярные клетки, возможно как оплодотворение сперматозоидами разных видов, так и самоплодотворение.
Предотвращение полиспермии
Большинство животных имеют моноспермические яйца, что означает проникновение только одного сперматозоида. Некоторые яйца защищены от проникновения более одного сперматозоида благодаря свойству их поверхности, в то время как другие обеспечивают эту функцию посредством оболочек. Способность некоторых яиц предотвращать полиспермию зависит от молекулярных изменений на их поверхности, происходящих в процессе созревания (оогенеза). Незрелые яйца морских ежей могут связываться со сперматозоидами, но допускают проникновение нескольких из них, что означает неспособность к предотвращению полиспермии. Поскольку зрелые яйца большинства животных оплодотворяются до завершения мейоза и могут предотвращать полиспормию, специфические свойства поверхности яйца формируются к моменту остановки мейоза, когда яйцо готово к оплодотворению.
У некоторых млекопитающих защита от полиспермии зависит от свойств zona pellucida, которая предотвращает проникновение дополнительных сперматозоидов после начала движения первого через zona (так называемая реакция zona). У других млекопитающих эта реакция либо ослаблена, либо отсутствует, что проявляется в наличии множества сперматозоидов в пространстве между zona и поверхностью яйца. В таких случаях реакция предотвращения полиспермии происходит непосредственно на поверхности яйца. Яйца некоторых животных (в частности, амфибий, птиц, рептилий и акул) от природы полиспермичны, однако только одно ядро сперматозоида сливается с ядром яйца, образуя ядро зиготы; оставшиеся сперматозоиды деградируют.
Формирование мембраны оплодотворения
Наиболее заметные изменения после оплодотворения наблюдаются на поверхности яйца. Хорошо известный пример - яйцо морского ежа - описан ниже. Первой реакцией на оплодотворение является поднятие вителлиновой мембраны с поверхности яйца. Изначально эта мембрана очень тонкая, но затем утолщается, приобретая структурированную молекулярную организацию и становится мембраной оплодотворения. Одновременно с этим происходит значительная перестройка молекулярной структуры поверхности яйца, и весь процесс занимает около одной минуты.
Когда сперматозоид прикрепляется к внешней поверхности яйца морского ежа, тонкая вителлиновая мембрана поднимается. Это позволяет мембранам кортикальных гранул соприкоснуться с внутренней стороной плазматической мембраны яйца и слиться с ней, после чего гранулы раскрываются, выпуская содержимое в пространство между поверхностью яйца и поднятой вителлиновой мембраной. Часть содержимого гранул взаимодействует с вителлиновой мембраной, формируя мембрану оплодотворения. Если процесс слияния содержимого гранул и вителлиновой мембраны нарушается, мембрана остаётся тонкой. Другие вещества из кортикальных гранул покрывают поверхность яйца, создавая прозрачный слой, называемый гиалиновым слоем, который важен для удержания вместе клеток (бластомеров) при делении яйца. Таким образом, плазматическая мембрана оплодотворённой яйцеклетки становится мозаичной структурой, включающей как участки исходной плазматической мембраны неоплодотворённой яйцеклетки, так и участки, сформированные из мембран кортикальных гранул. Образование мембраны оплодотворения сопровождается изменением электрического заряда плазматической мембраны, которое называется потенциалом оплодотворения, и одновременным выделением ионов калия; оба эти явления схожи с процессами в стимулированном нервном волокне. Оплодотворение также увеличивает проницаемость плазматической мембраны яйцеклетки для различных молекул за счёт активации определённого мембранного механизма.
Формирование ядра зиготы
После проникновения в цитоплазму яйцеклетки ядро сперматозоида, теперь называемое мужским пронуклеусом, начинает увеличиваться в размере, а его хромосомный материал рассеивается и становится схожим с женским пронуклеусом. Хотя мембранная оболочка мужского пронуклеуса быстро разрушается, вокруг него сразу формируется новая оболочка. Мужской пронуклеус, повернувшись на 180 градусов, движется к ядру яйца, сопровождаясь двумя структурами (центриолями), участвующими в делении клетки. После контакта мужского и женского пронуклеусов центриоли сперматозоида формируют первое веретено деления, предшествующее делению оплодотворённой яйцеклетки. В некоторых случаях слияние двух пронуклеусов может происходить посредством объединения мембран; при этом две близлежащие мембраны сливаются, образуя непрерывную оболочку, окружающую ядро зиготы.
Многие ранние исследования, посвященные биохимическим изменениям во время оплодотворения, сосредоточивались на метаболизме дыхания яйца. Однако результаты часто оказывались обманчивыми; например, яйцо морского ежа демонстрировало возросшее потребление кислорода сразу после оплодотворения или партеногенетической активации, что казалось подтверждением гипотезы о снятии метаболического блока в неоплодотворенном яйце. Тем не менее, сравнивая различные виды, оказалось, что повышение потребления кислорода у оплодотворенных яиц морского ежа не является универсальным правилом; на самом деле, у большинства животных яйцеклеток этот показатель не изменяется и может даже временно снижаться в момент оплодотворения.
В момент оплодотворения яйцеклетка содержит необходимые компоненты для синтеза белка и последующего развития на ранней стадии эмбрионального роста, называемой бластулой. Непосредственно после оплодотворения синтез белка управляется мессенджер-РНК, образованной в процессе оогенеза и хранящейся в яйцеклетке. К синтезу белка до стадии бластулы, а у млекопитающих и ранее, подключаются рибосомы, которые уже присутствуют в неоплодотворенном яйце; новые рибосомы и молекулы другой РНК, участвующей в синтезе белка, трансферной РНК, синтезируются на более поздней стадии эмбрионального развития (гаструляция). Опыт показал, что яйца, оплодотворенные и развивающиеся в присутствии актиномицина, ингибирующего синтез РНК, не только достигают стадии бластулы, но и имеют такую же скорость синтеза белка, как и необработанные эмбрионы.
У неоплодотворенных яиц морских ежей, как и у яиц других морских организмов, до сих пор исследованных, синтез белка происходит крайне медленно. Это указывает на то, что в неоплодотворенных яйцах присутствует одно или несколько веществ, препятствующих синтезу белков. После оплодотворения скорость синтеза белка резко увеличивается, что, вероятно, связано с устранением или изменением ингибитора. В яйцах морских ежей медленный синтез белка, по-видимому, обусловлен специфическими характеристиками рибосом. В неоплодотворенных яйцах большинство рибосом существуют в виде единичных рибосом (моносом), но вскоре после оплодотворения они начинают взаимодействовать с молекулами мРНК, образуя полирибосомы, которые являются активными единицами для синтеза белка. Аналогичные процессы происходят и в яйцах других морских организмов. Низкая эффективность синтеза белка в рибосомах неоплодотворенных яиц морского ежа вызвана наличием ингибитора, который препятствует связыванию мРНК с рибосомами. Этот ингибитор удаляется почти сразу после оплодотворения, вероятно, посредством ферментативного распада.
Таким образом, в яйце морских ежей общая эффективность синтеза белка регулируется на уровне рибосом, и первой стадией активации синтеза белка после оплодотворения является "активация" рибосом.
У позвоночных, таких как амфибии, активация синтеза белка начинается после завершения мейоза и овуляции яйцеклетки. В этом случае синтез белка запускается посредством связывания мРНК ооцита с рибосомами.
Ставьте лайк и подписывайтесь на наш канал, чтобы больше получать полезной информации.